HTTP 200 OK
Allow: GET, HEAD, OPTIONS
Content-Type: application/json
Vary: Accept
{
"count": 319,
"next": null,
"previous": "https://jabet.bsmiab.org/articles/?format=api&page=31",
"results": [
{
"id": 22,
"slug": "178-1519798391-expression-patterns-of-the-phosphoproteins-and-total-proteins-in-tlqp-21-a-vgf-derived-peptide-treated-sh-sy5y-cells",
"featured": false,
"slider": false,
"issue": "Vol1 Issue2",
"type": "original_article",
"manuscript_id": "178-1519798391",
"recieved": "2018-03-04",
"revised": null,
"accepted": "2018-03-23",
"published": "2018-05-08",
"pdf_file": "https://jabet.bsmiab.org/media/pdf_file/2023/09/178-1519798391.pdf",
"title": "Expression patterns of the phosphoproteins and total proteins in TLQP-21 (a VGF derived peptide) treated SH-SY5Y cells",
"abstract": "<p>VGF (non-acronymic), belonging to a large granin family, gives rise to a number of bioactive peptides by proteolysis and exert an extensive array of biological effects on energy metabolism, pain modulation, gastric secretion function, reproduction, mood regulation, and, diabetes. Among VGF-derived peptides, TLQP-21 (The first four amino acids, in short TLQP (Thr-Leu-Gln-Pro) generalizes the nomenclature of the peptide by its length) is the most studied although little is known yet about downstream molecular mechanisms of action of VGF-derived peptides like TLQP-21. So here as a preliminary analysis, total protein expression was carried out in addition to the phosphoproteomic study of SH-SY5Y cells treated with TLQP-21, using the same cell extracts. Comparison of simple 1D SDS-PAGE gels stained with SYPRO® Ruby protein gel stain was carried out to assess whether changes in protein expression could be seen even at such low separation resolution. Expression of several proteins most likely Microtubule-associated protein 1B (MW 271 kDa), Tubulin beta chain (MW 57), Tubulin beta-4B chain (MW 50), Alpha-2-macroglobulin (MW 163), etc. in TLQP-21 treated and control samples was found significantly different, indicating that the peptide TLQP-21 exerts biological effects on SH-SY5Y cells. Further studies are required to validate the identity of the modulated proteins, obtained from mass spectrometry. Identification of modulated proteins after TLQP-21 treatment would open new avenues to discover the molecular mechanisms of its physiological and pharmacological state.</p>",
"journal_reference": "J Adv Biotechnol Exp Ther. 2018; 1(2) : 43-48",
"academic_editor": "Dr. Masud Parvez, University Of Toronto, Canada.",
"cite_info": "Akhter MH, Requena JR.Expression patterns of the phosphoproteins and total proteins in TLQP-21 (a VGF derived peptide) treated SH-SY5Y cells. J Adv Biotechnol Exp Ther. 2018; 1(2) : 43-48.",
"keywords": [
"TLQP-21",
"SH-SY5Y",
"VGF",
"Gel stain",
"Phosphoprotein"
],
"DOI": "10.5455/jabet.2018.d8",
"sections": [
{
"section_number": 1,
"section_title": "INTRODUCTION",
"body": "<p>Protein phosphorylation provides key informations within signal transduction cascades and protein function modulations. It plays a key role in the regulation of most of the aspects of cellular biology [<a href=\"#r-1\">1</a>]. To detect protein phosphorylation directly in polyacrylamide gels, fluorescence-based detection technology, known as phosphoprotein gel stain, has been introduced [<a href=\"#r-2\">2</a>, <a href=\"#r-3\">3</a>] without using any antibody or radioisotope, compatible with mass spectrometry [<a href=\"#r-1\">1</a>]. For phosphoproteomic study as well as for identification of kinase targets in signaling pathways, phosphoprotein gel stain is a suitable method [<a href=\"#r-1\">1</a>].<br />\r\nSYPRO® Ruby dye is a quantitative, total-protein stain. Determining the ratio of Pro-Q® Diamond dye to SYPRO® Ruby dye signal intensities for each band or spot provides a measure of the phosphorylation level normalized to the total amount of protein. Using both stains in combination, it is possible to distinguish a lightly phosphorylated, high-abundance protein from a heavily phosphorylated, low-abundance protein. Thus, analysis of (phosphor and/or total) proteins by Pro-Q® Diamond phosphoprotein gel stain becomes most useful as well as most effective when used in combination with SYPRO® Ruby protein gel stain.<br />\r\nVGF (a non-acronymic name) is not to be confused with VEGF (vascular endothelial growth factor). The VGF gene was first recognized as a nerve growth factor (NGF) responsive gene. NGF33.1, a nervous system-specific mRNA was cloned by treatment of PC12 cells with NGF. For the first time, in 1985 Levi et al. [<a href=\"#r-4\">4</a>] designated the clone corresponding to the NGF-inducible mRNA as VGF, after successful clarification of the nucleic acid as well as amino acid sequences of the NGF33.1 cDNA clone. The term ‘VGF’ was coined very interestingly as the selection of this clone was from plate V of the nerve Growth Factor induced PC12 cell cDNA library [<a href=\"#r-4\">4</a>, <a href=\"#r-5\">5</a>].<br />\r\nFrom VGF, several peptides are derived like APPG-40, APPG-37, GRPE-37, NERP-1, NERP-2, NAPP-129 (VGF 20), VGF 18, HFHH-51 (VGF 6), HHPD-1, AQEE-30 (Peptide V), LQEQ-19, TLQP-21 and TLQP-62 (VGF 10). Out of these peptides, TLQP-21 is of great importance because of its several physiological roles. TLQP-21 plays vital roles in the following physiological actions: Energy expenditure [<a href=\"#r-6\">6</a>, <a href=\"#r-7\">7</a>, <a href=\"#r-8\">8</a>] metabolic functions [<a href=\"#r-6\">6</a>, <a href=\"#r-15\">15</a>], glucose-stimulated insulin secretion (GSIS) [<a href=\"#r-9\">9</a>], nociception [<a href=\"#r-10\">10</a>, <a href=\"#r-11\">11</a>, <a href=\"#r-12\">12</a>], blood pressure/hypertension regulation [<a href=\"#r-13\">13</a>], gastric contractility [<a href=\"#r-14\">14</a>, <a href=\"#r-15\">15</a>], regulation of gastric acid secretion [<a href=\"#r-16\">16</a>,<a href=\"#r-17\"> 17</a>, <a href=\"#r-18\">18</a>], reproduction [<a href=\"#r-19\">19</a>, <a href=\"#r-20\">20</a>], stress [<a href=\"#r-21\">21</a>, <a href=\"#r-22\">22</a>], neuroprotective agent [<a href=\"#r-23\">23</a>], anorexia [<a href=\"#r-6\">6</a>, <a href=\"#r-7\">7</a>]. To the best of our knowledge, no proteomic or phosphoproteomic studies have yet focused on the effect of VGF derived bioactive peptide TLQP-21 on signaling pathways in SH-SY5Y cells.<br />\r\nTaken together all these in considerations, a study on the expression of phosphoproteins and total proteins in TLQP-21 treated SH-SY5Y cells was conducted to conclude whether modulation in protein expression could be found using Pro-Q® Diamond phosphoprotein gel stain, in conjunction with SYPRO® Ruby protein gel stain.</p>"
},
{
"section_number": 2,
"section_title": "MATERIALS AND METHODS",
"body": "<p><strong>TLQP-21</strong><br />\r\nTLQP-21 (human, molecular weight 2490.88 Da) and modified TLQP-21 (human) containing biotin at N-terminus and a cysteine residue at the C-terminus with total molecular weight 2820.32 Da, were purchased from ChinaPeptides Co. Ltd., Shanghai. The purity >95% of the both peptides, TLQP-21 and biotinlyted TLQP-21 were confirmed by HPLC and MS analysis. It was in the form of white lyophilized powder and stored at -21˚C immediately upon arrival for short time storage and at -80˚C for long term storage, as per instruction of the supplier. The peptide sequence is – TLQPPSALRRRHYHHALPPSR<br />\r\nThr – Leu – Gln – Pro – Pro – Ser – Ala – Leu – Arg – Arg – Arg – His – Tyr -His – His – Ala – Leu – Pro – Pro – Ser – Arg<br />\r\nTLQP-21 solutions were constituted by dissolving the lyophilized powder in filtered (0.22 μm, Millex, Merck Millipore Ltd.) PBS, and were used instantly or was kept at -80 ˚C for long term storage.</p>\r\n\r\n<p><br />\r\n<strong>SH-SY5Y cell culture</strong><br />\r\nSH-SY5Y (European Collection of Cell Cultures, ECACC; catalog number-94030304) is a thrice cloned (SK-N-SH → SH-SY → SH-SY5 → SH-SY5Y) subline of the neuroblastoma cell line SK-N-SH which was established in 1970 from a metastatic bone tumor of a four year-old female with neuroblastoma [<a href=\"#r-24\">24</a>, <a href=\"#r-25\">25</a>]. As per standard protocol, the cells were always used at less than 20 passages and were grown in 100×20 mm Falcon Petri dishes (Life Sciences) on a culture medium composed of 1:1 F12 HAM (Sigma Aldrich) and Earle’s Balanced Salt Solution (EBSS) (Sigma Aldrich), which was supplemented with 1% Penicilin-Streptomicin (P/S) (Invitrogen), 15% fetal bovine serum (FBS) (Gibco), 1% Non-Essential Amino Acids (NEAA) (Sigma Aldrich) and 1% Glutamine (Gln) (Sigma Aldrich). And for optimum growth, 5% CO2-humidified incubator was kept at 37 ˚C.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Phosphoprotein gel staining</strong><br />\r\nSH-SY5Y cells in Petri dishes with confluent growth were incubated for 6 hours with the peptide TLQP-21 at a concentration of 1μg/ml. Then the cell homogenates were prepared as discussed above. As control, cells without peptide incubation were grown, then homogenized under the same conditions as treated cells.<br />\r\nCell homogenate samples (20 μl) were boiled in 2X Laemmli buffer (Bio-Rad) for 10 minutes at 100 ˚C, spun, and loaded to SDS-PAGE gel. After electrophoresis (200V, 01h), the gels were washed in dH2O for 10 minutes. Then the gels were kept with fixing solution composed of 50% methanol and 10% acetic acid in an orbital shaker step at 50 rpm for half an hour at room temperature. The fixation step was repeated once. Followed by washing the gel with water three times each for ten minutes, the gels were incubated for one and half hour with Pro-QR Diamond (Invitrogen) gel stain in the dark with agitation at 50 rpm. The gels were then destained with Pro-QR Diamond phosphoprotein gel destaining solution (20 % acetonitrile, 50 mM sodium acetate, pH 4) for 30 minutes with shaking. The destaining procedure was repeated two times more, followed by washing the gels two times each for 5 minutes with water. And then the gel was imaged using a Typhoon FLA9500 scanner at a 100 μm resolution at 532nm green laser.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>For SYPRO® Ruby protein gel staining</strong><br />\r\nSamples (25 μl) from cell homogenates of the cells treated with the peptide and not treated were heated in 2X Laemmli buffer (Bio-Rad) for 10 minutes at 100 °C, spun, and loaded to SDS-PAGE gel. After electrophoresis (200V, 01h), the gel was taken out from cassette, followed by continuous washing for 10 minutes in dH2O. Then the gel was treated for 60 minutes at room temperature with fixing solution, consists of 10% methanol and 7% acetic acid in an orbital shaker step at 50 rpm, followed by overnight incubation with SYPRO® Ruby Protein Gel Stain at room temperature with shaking. The gel was then placed into a staining container covering with a lid to protect it from the light, in addition, the container was wrapped in aluminum foil to further shield the stain from light during the staining process, as per instruction of the supplier. The gel was shifted to a clean staining dish followed by staining for overnight. Then the gel was washed with the fixing solution in the same condition of staining followed by 5 minutes washing in dH2O. Finally, the gel was taken from the container to take the image in a MolecularImager® Gel DocTM system (Bio Rad, Hercules). To capture the best image, the highest sensitivity of the CCD camera was used at a resolution of 1392 x 1040 pixels with 12 bit gray scale levels per pixel.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Protein detection using mass spectrometric analysis</strong><br />\r\nFrom the gels, spots were chosen for mass spectrometric analysis and were excised with a pipette tip and then in-gel digestion was performed manually with trypsin following the standard protocol described by Shevchenko et al., 2006. The excised spots were washed three times with 100 μl of ammonium bicarbonate 50 mM in 50% methanol (grade HPLC Scharlau) and reduced with 10 mM DTT (SERVA Electrophoresis GmbH). After this, the gel pieces were washed three times with ammonium bicarbonate, dried in a SpeedVac (Thermo Scientific) and alkylated with 55 mM iodoacetamide (IAA) (Sigma- Aldrich). The gel pieces were washed again with ammonium bicarbonate, dehydrated with acetonitrile and dried again in a SpeedVac (Thermo Scientific). Trypsin, modified with porcine (Promega) was added in 20 mM ammonium bicarbonate at the final concentration of 20 ng/mL and digestion let proceed overnight at 37ºC. Peptide extraction from the gel pieces was carried out three times with 40 μL of 60% acetonitrile in 0.5% of formic acid. Then the extracts were collected followed by drying in the SpeedVac (Thermo Scientific) and kept it at -20ºC. After digestion the spots were identified using a model 4800 MALDITOF/TOF mass spectrometer (ABSciex, Framingham, MA, EEUU) at the Proteomics Lab of the Foundation IDICHUS (University Clinical Hospital, University of Santiago de Compostela, Santiago de Compostela, Spain), as per protocol described above. Programmed laboratory analysis of mass data was conducted by using the 4000 Series. It is to be noted that to get the best results, all of the MS/MS spectra were performed taking the considerations of metastable suppression as well as by selecting the precursor ions with a relative resolution of 300 (FWHM).<br />\r\nFor convenience, through the GPS Explorer Software v3.6 both of the MS/MS spectra data and Explorer Software V3.5. MS data were pooled together; followed by the database search which was performed with the Mascot v2.1 search tool (Matrix Science, London, UK) screening SwissProt (release 56.0). It is noteworthy that carbamidomethyl cysteine was set as a fixed modification and oxidized methionine was set as potential variable modification, following by searches which were restricted to human taxonomy. The precursor mass tolerance was fixed at 30 ppm whereas the MS/MS tolerance was set at 0.35 Da. And 1 missed tryptic cleavage site was allowed. All spectra and database findings were manually inspected and afterwards all these were detailed using the softwares, as mentioned above [<a href=\"#r-27\">27</a>, <a href=\"#r-28\">28</a>, <a href=\"#r-29\">29</a>].</p>"
},
{
"section_number": 3,
"section_title": "RESULTS",
"body": "<p>As supplementary confirmation that TLQP-21exerts biological effects in the model system, SH-SY5Y cells; homogenates from TLQP-21 treated or not treated (control) cells were used to conduct 1D SDS-PAGE, then it was stained with the dye Pro-Q Diamond, followed by SYPRO® Ruby dye. As seen in the <a href=\"#figure1\">Figure 1</a>, at position 3, 4 and 5; band intensity becomes higher in SYPRO® Ruby dye staining indicate the nonphosphorylated proteins. Differences in band intensity were seen at position 1 to 5 of the gels. The bands’ intensity at 1 (A, Pro Q® Diamond) and 3 (both in A, Pro Q® Diamond and B, SYPRO® Ruby dye) were less in the samples treated with the peptide in comparison to not treated (control) ones suggesting that the peptide on SH-SY5Ycells might provoke dephosphorylation of specific phosphoproteins. The bands at position 1 and 3 from A, Pro Q® Diamond stained gel was cut and analysed by mass spectrometry, results are detailed in the <a href=\"#Table-1\">Table 1</a>.</p>\r\n\r\n<div id=\"figure1\">\r\n<figure class=\"image\"><img alt=\"\" height=\"416\" src=\"/media/article_images/2024/16/09/178-1519798391-Figure1.jpg\" width=\"492\" />\r\n<figcaption><strong>Figure 1. </strong>Comparative selectivity of A). Pro-Q® Diamond phosphoprotein staining and B). SYPRO® Ruby (total) protein staining. Here the gel, at first, was stained with Pro-Q Diamond dye, then the same gel was subjected to post staining with SYPRO Ruby protein gel stain to picture out of all the proteins of the cell homogenates treated (T) and control (C) with the peptide showing differences in protein expression at position 1, 2, 3, 4 and 5.</figcaption>\r\n</figure>\r\n</div>\r\n\r\n<p>A comparison was made among Pro Q® Diamond staining, SYPRO® Ruby protein gel stain and simple 1D SDS-PAGE gels: Several bands with altered intensity in the SYPRO® Ruby experiment were of the same positions in the gels as those in the Pro Q® Diamond, suggesting that both techniques were detecting, to some extent, similar changes. This also suggested that the proteins with altered phosphorylation status were probably abundant proteins, hence their changes were “picked up” by SYPRO® Ruby staining.</p>\r\n\r\n<p>Out of the proteins listed in the <a href=\"#Table-1\">table 1</a>, several proteins like Microtubule-associated protein 1B (MW 271 kDa), Tubulin beta chain (MW 57), Tubulin beta-4B chain (MW 50), Alpha-2-macroglobulin (MW 163) were of interests, as these are related with neurological and other functions of VGF derived peptide TLQP-21 [<a href=\"#r-27\">27</a>,<a href=\"#r-29\"> 29</a>]. Further studies are required to validate the results obtained from mass spectrometry and to illustrate the further downstream consequences with reference to clinical studies.</p>\r\n\r\n<div id=\"Table-1\">\r\n<p><strong><a href=\"https://jabet.bsmiab.org/table/178-1519798391-table1/\">Table 1.</a> Table 1. </strong>List of proteins with altered expression levels in Pro Q® Diamond stained gel.</p>\r\n</div>"
},
{
"section_number": 4,
"section_title": "DISCUSSION",
"body": "<p>In TLQP-21 treated rat pituitary tumor cell lines (GH3), no difference was found in pERK and pAMPK though there was difference in expression of pAKT and p-p38 [<a href=\"#r-26\">26</a>]. In another study, in rat cerebellar granule cells (CGCs) TLQP-21 was shown to activate ERK½ significantly. Akt phosphorylation was found to be increased after 15 minute of treatment with TLQP-21, while using insulin-like growth factor-1 (IGF-1) phosphorylation of Akt increased further after the same time interval. IGF-1 treatment augmented Akt phosphorylation after 24 and 48 h of incubation whereas TLQP-21 did not modify the amount of phosphorylated Akt, although it is to mentioned here that total Akt and α-tubulin were found to be expressed more than before in both cases in rat CGCs [<a href=\"#r-22\">22</a>].</p>\r\n\r\n<p>In another study of TLQP-21 induced signal transduction pathway in mice 3T3-L1 adipocytes, TLQP-21 did not bring any change in expression of Akt (Ser473), PKC (protein kinase C; pan Ser660), p38 (Thr180/Tyr182)and JNK (c-Jun N-terminal kinase; Thr183/Tyr185, PKA and HSL , whereas TLQP-21 increased phosphorylation of AMPK and ERK [<a href=\"#r-6\">6</a>].<br />\r\nFrom literature review it is evident that this is the first study in human cell line, to observe the effect of the peptide, TLQP-21 whether it modulates the total proteins or the phosphoproteins or not. The significance of the study lies into the fact that it highlights a vital opening point for more advanced exploration into cell signaling by TLQP-21. Further proteomic analysis, followed by 2D Gel- or LC-MALDI TOF/TOF will help us for better understanding in this regard.</p>"
},
{
"section_number": 5,
"section_title": "ACKNOWLEDGMENTS",
"body": "<p>Md. Shamim Akhter is the recipient of an Erasmus Mundus EXPERTS – II scholarship for doctoral program.</p>"
},
{
"section_number": 6,
"section_title": "CONFLICT OF INTEREST",
"body": "<p>The author declares that no conflict of interest exists.</p>"
}
],
"figures": [
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/16/09/178-1519798391-Figure1.jpg",
"caption": "Figure 1: Comparative selectivity of A). Pro-Q® Diamond phosphoprotein staining and B). SYPRO® Ruby (total) protein staining. Here the gel, at first, was stained with Pro-Q Diamond dye, then the same gel was subjected to post staining with SYPRO Ruby protein gel stain to picture out of all the proteins of the cell homogenates treated (T) and control (C) with the peptide showing differences in protein expression at position 1, 2, 3, 4 and 5.",
"featured": false
}
],
"authors": [
{
"id": 40,
"affiliation": [
{
"affiliation": "Biotechnology and Genetic Engineering Discipline, Khulna University, Khulna-9208, Bangladesh."
}
],
"first_name": "Md. Shamim",
"family_name": "Akhter",
"email": "shamim11akhter@gmail.com",
"author_order": 1,
"ORCID": "https://orcid.org/0000-0001-5630-3906",
"corresponding": true,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "Dr. Md. Shamim Akhter, Biotechnology and Genetic Engineering Discipline, Khulna University, Khulna,\r\nBangladesh. E-mil: shamim11akhter@gmail.com",
"article": 22
},
{
"id": 41,
"affiliation": [
{
"affiliation": "CIMUS Biomedical Research Institute, University of Santiago de Compostela-IDIS, Santiago de Compostela-15782, Spain."
}
],
"first_name": "Jesús R.",
"family_name": "Requena",
"email": null,
"author_order": 2,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 22
}
],
"views": 534,
"downloads": 77,
"references": [
{
"id": 237,
"serial_number": 1,
"pmc": null,
"reference": "Schulenberg B, Goodman TN, Aggeler R, Capaldi RA, Patton WF. Characterization of dynamic and steady-state protein phosphorylation using a fluorescent phosphoprotein gel stain and mass spectrometry. Electrophoresis 2004; 25 : 2526-2532.",
"DOI": null,
"article": 22
},
{
"id": 238,
"serial_number": 2,
"pmc": null,
"reference": "Steinberg T, Agnew B, Gee K, Leung W, Goodman T, Schulenberg B, et al. Proteomics 2003; 3: 1128–1144.",
"DOI": null,
"article": 22
},
{
"id": 239,
"serial_number": 3,
"pmc": null,
"reference": "Martin K, Steinberg T, Cooley L, Gee K, Beechem J, Patton W. Proteomics 2003; 3: 1244–1255.",
"DOI": null,
"article": 22
},
{
"id": 240,
"serial_number": 4,
"pmc": null,
"reference": "Levi A, Eldridge JD., Paterson BM. Molecular cloning of a gene sequence regulated by nerve growth factor. Science 1985; 229 (4711):393-395.",
"DOI": null,
"article": 22
},
{
"id": 241,
"serial_number": 5,
"pmc": null,
"reference": "Possenti R, Eldridge JD., Paterson BM., Grasso A, Levi A. A protein induced by NGF in PC12 cells is stored in secretory vesicles and released through the regulated pathway. EMBO J 1989; 8(8): 2217-2223.",
"DOI": null,
"article": 22
},
{
"id": 242,
"serial_number": 6,
"pmc": null,
"reference": "Possenti R, Muccioli G, Petrocchi P, Cero C, Cabassi A, Vulchanova L. et al. Characterization of a novel peripheral pro-lipolytic mechanism in mice: role of VGF-derived peptide TLQP-21. Biochem J 2012; 441 (1): 511-22.",
"DOI": null,
"article": 22
},
{
"id": 243,
"serial_number": 7,
"pmc": null,
"reference": "Jethwa PH, Warner A, Nilaweera KN, Brameld JM, Keyte JW, Carter, WG et al. VGF-derived peptide, TLQP-21, regulates food intake and body weight in Siberian hamsters. Endocrinology 2007; 148: 4044-4055.",
"DOI": null,
"article": 22
},
{
"id": 244,
"serial_number": 8,
"pmc": null,
"reference": "Bartolomucci A, Corte GL, Possenti R, Locatelli V, Rigamonti AE, Torsello A, et al. TLQP-21, a VGF-derived peptide, increases energy expenditure and prevents the early phase of diet-induced obesity. Proc. Natl. Acad. Sci. U. S. A. 2006; 103 : 14584–14589.",
"DOI": null,
"article": 22
},
{
"id": 245,
"serial_number": 9,
"pmc": null,
"reference": "Stephens SB, Schisler JC, Hohmeier HE, An J, Sun AY, Pitt GS, et al. A VGF-derived peptide attenuates development of type 2 diabetes via enhancement of islet β-cell survival and function. Cell Metab 2012; 16 (1): 33-43. doi: 10.1016/j.cmet.2012.05.011.",
"DOI": null,
"article": 22
},
{
"id": 246,
"serial_number": 10,
"pmc": null,
"reference": "Fairbanks CA, Peterson CD, Speltz RH, Riedl, MS, Kitto, KF, Dykstra, JA, et al. The VGF-derived peptide TLQP-21 contributes to inflammatory and nerve injury induced hypersensitivity. Pain 2014; 155: 1229–1237.",
"DOI": null,
"article": 22
},
{
"id": 247,
"serial_number": 11,
"pmc": null,
"reference": "Chen YC, Pristerá A, Ayub M, Swanwick RS., Karu K, Hamada Y, et al. Identification of a receptor for neuropeptide VGF and its role in neuropathic pain. J Biol Chem 2013; 288 (48): 34638-46.",
"DOI": null,
"article": 22
},
{
"id": 248,
"serial_number": 12,
"pmc": null,
"reference": "Rizzi R, Bartolomucci A, Moles A, D’Amato F, Sacerdote P, Levi A, et al. The VGF-derived peptide TLQP-21: a new modulatorypeptide for inflammatory pain. Neurosci Lett 2008; 441: 129–133.",
"DOI": null,
"article": 22
},
{
"id": 249,
"serial_number": 13,
"pmc": null,
"reference": "Fargali S, Garcia AL, Sadahiro M, Jiang C, Janssen WG, Lin WJ, et al. The granin VGF promotes genesis of secretory vesicles, and regulates circulating catecholamine levels and blood pressure. FASEB J 2014; 28: 2120–2133.",
"DOI": null,
"article": 22
},
{
"id": 250,
"serial_number": 14,
"pmc": null,
"reference": "Severini C, La Corte G, Improta G, Broccardo M, Agostini S, Petrella C, et al. In vitro and in vivo pharmacological role of TLQP-21, a VGF-derived peptide, in the regulation of rat gastric motor functions. British journal of pharmacology 2009; 157: 984-993.",
"DOI": null,
"article": 22
},
{
"id": 251,
"serial_number": 15,
"pmc": null,
"reference": "Bartolomucci A, Moles A, Levi A, Possenti R. Pathophysiological role of TLQP-21: gastrointestinal and metabolic functions. Eat Weight Disord. 2008; 13(3): e49-54. PMID: 19011364.",
"DOI": null,
"article": 22
},
{
"id": 252,
"serial_number": 16,
"pmc": null,
"reference": "Sibilia V, Pagani F, Bulgarelli I, Tulipano G, Possenti R, Guidobono F. Characterization of the mechanisms involved in the gastric antisecretory effect of TLQP-21, a vgf-derived peptide, in rats. Amino Acids 2012; 42 (4): 1261-8. doi: 10.1007/s00726-010-0818-6. Epub 2010 Dec 4.",
"DOI": null,
"article": 22
},
{
"id": 253,
"serial_number": 17,
"pmc": null,
"reference": "Sibilia V, Pagani F, Bulgarelli I, Mrak E, Broccardo M, Improta G, et al. TLQP-21, a VGF-derived peptide, prevents ethanol-induced gastric lesions: insights into its mode of action. Neuroendocrinology 2010a; 92 (3): 189-97. doi: 10.1159/000319791.",
"DOI": null,
"article": 22
},
{
"id": 254,
"serial_number": 18,
"pmc": null,
"reference": "Sibilia V, Pagani F, Bulgarelli I, Tulipano G, Possenti R, Guidobono F. Characterization of the mechanisms involved in the gastric antisecretory effect of TLQP-21, a VGF-derived peptide, in rats. Amino Acid 2010b. 10.1007/s00726-010-0818-6.",
"DOI": null,
"article": 22
},
{
"id": 255,
"serial_number": 19,
"pmc": null,
"reference": "Aguilar E, Pineda R, Gayta´ n F, Sa´nchez-Garrido MA, Romero M, Romero-Ruiz A, et al. Characterization of the reproductive effects of the VGF-derived peptide TLQP-21 in female rats: in vivo and in vitro studies. Neuroendocrinology 2013; 98: 38–50.",
"DOI": null,
"article": 22
},
{
"id": 256,
"serial_number": 20,
"pmc": null,
"reference": "Pinilla L, Pineda R, Gaytan F, Romero M, Garcia-Galiano D, Sanchez-Garrido M.A, et al. Characterization of the reproductive effects of the anorexigenic VGF-derived peptide TLQP-21: in vivo and in vitro studies in male rats. American Journal of Physiology Endocrinology and Metabolism 2011; 300: 837-847.",
"DOI": null,
"article": 22
},
{
"id": 257,
"serial_number": 21,
"pmc": null,
"reference": "Razzoli M, Bo E, Pascucci T, Pavone F, D’Amato FR, Cero, et al. Implication of the VGF-derived peptide TLQP-21 in mouse acute and chronicstress responses. Behav. Brain Res 2012; 229: 333–339.",
"DOI": null,
"article": 22
},
{
"id": 258,
"serial_number": 22,
"pmc": null,
"reference": "Bartolomucci A, Possenti R, Mahata SK., Fischer-Colbrie R, Loh YP, Salton SR. The extended graninfamily: structure, function, and biomedical implications. Endocr Rev 2011; 32 (6): 755-97.",
"DOI": null,
"article": 22
},
{
"id": 259,
"serial_number": 23,
"pmc": null,
"reference": "Severini C, Ciotti MT, Biondini L, Quaresima S, Rinaldi AM, Levi A, Frank C, Possenti R. TLQP-21, a neuroendocrine VGF-derived peptide, prevents cerebellar granule cells death induced by serum and potassium deprivation. J Neurochem 2008; 104: 534-544.",
"DOI": null,
"article": 22
},
{
"id": 260,
"serial_number": 24,
"pmc": null,
"reference": "Biedler JL., Helson L, Spengler BA. Morphology and growth, tumorigenicity, and cytogenetics of human neuroblastoma cells in continuous culture. Cancer Res. 1973; 33 (11): 2643–52.",
"DOI": null,
"article": 22
},
{
"id": 261,
"serial_number": 25,
"pmc": null,
"reference": "Biedler JL, Roffler-Tarlov S, Schachner M, Freedman LS. Multiple neurotransmitter synthesis by human neuroblastoma cell lines and clones. Cancer Res 1978; 38 : 3751–7.",
"DOI": null,
"article": 22
},
{
"id": 262,
"serial_number": 26,
"pmc": null,
"reference": "Passeri PP, Biondini L, Mongiardi MP, Mordini N, Quaresima S, Frank C, et al. Neuropeptide TLQP-21, a VGF internal fragment, modulates hormonal gene expression and secretion in GH3 cell line. Neuroendocrinology 2013; 97: 212–224. DOI: 10.1159/000339855.",
"DOI": null,
"article": 22
},
{
"id": 263,
"serial_number": 27,
"pmc": null,
"reference": "Akhter S, Chakraborty S, Moutinho,D, AÂlvarez-Coiradas E, Rosa I, Viñuela J, et al. The human VGF-derived bioactive peptide TLQP-21 binds heat shock 71 kDa protein 8 (HSPA8) on the surface of SH-SY5Y cells. PLoS ONE 2017; 12(9): e0185176. https://doi.org/10.1371/journal. pone.0185176.",
"DOI": null,
"article": 22
},
{
"id": 264,
"serial_number": 28,
"pmc": null,
"reference": "Ayub M. Investigating the mechanisms of action of VGF-derived peptides in the nervous system. Ph D Thesis. 2012; Imperial College, London.",
"DOI": null,
"article": 22
},
{
"id": 265,
"serial_number": 29,
"pmc": null,
"reference": "Akhter S. Isolation of VGF derived neuropeptide receptor. Ph. D. Thesis. 2015; University of Santiago de Compostela, Spain.",
"DOI": null,
"article": 22
}
]
},
{
"id": 20,
"slug": "178-1524741069-long-term-administration-of-gentamicin-affects-hemato-biochemical-parameters-and-liver-architecture-of-swiss-albino-mice",
"featured": false,
"slider": false,
"issue": "Vol1 Issue2",
"type": "original_article",
"manuscript_id": "178-1524741069",
"recieved": "2018-02-12",
"revised": null,
"accepted": "2018-03-12",
"published": "2018-05-08",
"pdf_file": "https://jabet.bsmiab.org/media/pdf_file/2023/03/178-1524741069.pdf",
"title": "Long term administration of gentamicin affects hemato-biochemical parameters and liver architecture of Swiss Albino mice",
"abstract": "<p>Gentamicin is most frequently used aminoglycoside antibiotic. Despite its wide use, the effects of gentamicin have not been clearly studied in relation to alteration of hemato-biochemical parameters and liver injury. In the present study, to evaluate the effects of gentamicin on behavioral, hematological, biochemical and morphological parameters of liver, Swiss albino mice were divided into 4 experimental groups (group A: control; group B: pharmacological dose; group C: pharmacological dose rate with chronic treatment, and group D: high dose with chronic treatment). All the mice from group C and D showed dullness, fearness, roughness of the body coat, anorexia and weakness. Liver weight and size were increased significantly in the mice of group C and D than that of group A or group B. Similarly, in hematological study, Total erythrocyte count (TEC), Total leukocyte count (TLC) and hemoglobin (Hb) % values were decreased significantly, whereas, Alanine amino transferase (ALT) values were increased significantly in the mice of group C and D. In addition, congestion and dark coloration with hepatomegaly were found in treated group C and D. Histological study revealed that the liver parenchyma showed central vein congestion, lymphocytic infiltration, irregular size of hepatocyte and dilatation of sinusoids of treated group C and D. Taken together, our current study suggests that although pharmacological dose of Gentamicin has no adverse effect on liver but chronic pharmacological dose or chronic high dose has serious adverse effect on liver. These observations lead us to postulate that gentamicin induces liver tissue damage after long term treatment.</p>",
"journal_reference": "J Adv Biotechnol Exp Ther. 2018; 1(2) : 29-35",
"academic_editor": "Dr. Hasan-Al-Faruque, Daegu Gyeonbuk Institute of Science and Technology, South Korea.",
"cite_info": "Jannat N, Amin T, Sultana N, etal. Long term administration of gentamicin affects hemato-biochemical parameters and liver architecture of Swiss Albino mice. J Adv Biotechnol Exp Ther. 2018; 1(2) : 29-35.",
"keywords": [
"Liver",
"Mice",
"Gentamicin",
"Morphology",
"Hepatomegaly"
],
"DOI": "10.5455/jabet.2018.d6",
"sections": [
{
"section_number": 1,
"section_title": "INTRODUCTION",
"body": "<p>Liver is the largest gland in our body and plays most important functions not only in the storage and release of nutrients but also in the neutralization and elimination of a variety of toxic substances. Antibiotics are one of the most widely used choices of drugs. These drugs are used for prevention of many problems caused by infections. However, antibiotics have side effects and can damage various body organs including liver, kidney, brain, blood, skin, eyes and mouth [<a href=\"#r-1\">1</a>]. Among the antibiotics, gentamicin is the most widely studied aminoglycoside antibiotic used to treat severe infections of Gram-negative bacteria [<a href=\"#r-2\">2</a>]. The aminoglycoside antibiotic, gentamicin is synthesized by <em>Micromonospora purpurea</em> [<a href=\"#r-3\">3</a>]. The use of gentamicin has tremendously increased in human and veterinary practice due to their greater effectiveness against human, livestock and poultry diseases [<a href=\"#r-4\">4</a>]. But sometimes people of Bangladesh, particularly rural people are not concerned about many legal issues due to limited literacy. They purchase antibiotics without any prescription from physician or even when the practice is not legal. For treatment purposes, they use overdose of antibiotic for a long time which may cause adverse effects in human beings. In rural part of Bangladesh, 95% of the people consume drugs without any prescription and purchase drugs from local pharmacies; only 8% of them consume drugs according to the prescription from physicians [<a href=\"#r-5\">5</a>].<br />\r\nGentamicin is a heat stable antibiotic that remains active even after autoclaving, thus making it useful in the preparation of certain microbiological growth media. The action on bacteria is bactericidal and gentamicin has increased activity at alkaline pH. In case of gentamicin, oral absorption is minimal and for systemic use gentamicin must be given by the parenteral route. Uptake is rapid after intramuscular injection and it has a serum half – life of 75-110 minutes [<a href=\"#r-6\">6</a>]. The distribution of aminoglycosides antibiotic after an IV injection is virtually complete within 1 hour. The penetration of aminoglycosides across membranous barriers by simple diffusion is very limited due to polycationic nature of these antibiotics. Therefore, very low concentrations of aminoglycosides are found in cerebrospinal fluids or respiratory secretions [<a href=\"#r-7\">7</a>].<br />\r\nGentamicin is mainly used for clinical practice. The most frequently reported side effects associated with gentamicin therapy are ototoxicity, nephrotoxicity and hepatotoxicity [<a href=\"#r-8\">8</a>]. These forms of toxicity occur more frequently in patients who experience prolonged exposure to serum gentamicin concentrations of greater than 2 mcg/mL [<a href=\"#r-9\">9</a>].<br />\r\nAfter the use of gentamicin in cells, increased production of Reactive Oxygen Species (ROS) is effective in inducing toxic impacts of this drug on the structure and function of tissues [<a href=\"#r-10\">10</a>]. Gentamicin enhanced the production of superoxide anion, hydrogen peroxide and hydroxyl radicals by mitochondria [<a href=\"#r-11\">11</a>]. Free radicals cause Peroxidation of phospholipids membrane, DNA strand breakage, protein denaturation. These effects induce changes in membrane fluidity, thus the membrane gets permeable even to molecules as large as enzymes [<a href=\"#r-12\">12</a>]. Additionally, blood chemical investigation was conducted for more elucidation of the effects of tissue damage which could be provoked by gentamicin. Therefore, the effects of gentamicin must take into account as problem relating to human beings, animals and birds. Despite its wide use, gentamicin has not been definitively linked to instances of clinically apparent liver injury, but in the present study we investigated morphological and hemato-biochemical effects of gentamicin on liver in mice.</p>"
},
{
"section_number": 2,
"section_title": "MATERIALS AND METHODS",
"body": "<p><strong>Chemicals</strong><br />\r\nGentaren 10% (Reneta, Bangladesh), 100 ml bottle is an aminoglycoside antibiotic preparation. Gentamicin commonly known as a broad-spectrum antibiotic was purchased from local market.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Animals and treatments</strong><br />\r\nThe experimental male Swiss albino mice were collected from International Center for Diarrheal Disease Research (icddr’b), Mohakhali, Dhaka, Bangladesh. All the mice were possessed good health and devoid of any external deformities certified by the registered veterinarian from icddr’b. After procurement, all the mice were kept under close observation in order to acclimatize to the new environment for a period of one week prior to commencement of the experiment. All mice were raised under confinement as an intensive system. All experimental protocols were approved by the Animal Welfare and Ethical Committee, Faculty of Veterinary Science, Bangladesh Agricultural University. All efforts were made to minimize the number of mice used and their sufferings. Twenty male mice, aged 5-6 weeks old weighing 25-30 g were used for this experiment. The rats were housed four per one plastic cage, maintained on a 12h light/dark cycle at a constant temperature (70-740F) and humidity (45-60%) and provided water and rodent pellets ad libitum. For each individual, under study a record sheet with full details of each parameter were maintained. For the experimental purpose, the mice were randomly divided into four groups: Group A (Control), without any treatment; Group B (5mg/kg for 7 days), pharmacological dose; Group C (5mg/kg for 30 days), pharmacological dose rate with chronic treatment; Group D (10mg/kg for 30 days), high dose rate with chronic treatment. Each group contained five mice. Body weights of all mice were recorded before starting the treatment. After administration of Gentamicin all the mice were kept under close observation for entire 35days (30 days of treatment period and 5 days of post treatment).</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Clinical examination</strong><br />\r\nDuring the tenure of the treatment, the animals were carefully observed for behavioral study.<br />\r\nBlood chemical analysis<br />\r\nEach animal was euthanized by using chloroform before 2ml of blood was taken in 5 ml disposable syringe by cardiac puncture for estimation of various blood chemical parameters such as- TEC, TLC, Hb%, ALT. The blood sample was allowed to stand for one hour and centrifuged at 3000 rpm for 15 minutes. Eppendrof tubes were used for collection of serum and stored in freeze at -200C. ALT was measured by using commercially available kits. The number of RBC and WBC were calculated accordingly (Number of RBC= Number of counted ×1000 and express the results in millions per cu. Mm; Number of WBC= Number of counted×50 and the result is expressed in thousand per cu. Mm).</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Gross and histology</strong><br />\r\nAfter sacrifice of each animal sequentially, liver was collected from each animal and examined for gross study. For gross study, color, weight and size of liver was taken into consideration.<br />\r\nFor histological observation, 5 mm pieces were collected from different side of liver and immersed in 10% formalin for 48 hours. Then, the sample was washed in 10% phosphate buffer solution for 3 hours, dehydration was done by passing the tissue in the ascending grade of alcohol, such as 70, 80, 90, 95, 100% (1), and 100% (2) each for 2 hour and finally 100% (3) for overnight, cleared in xylene and embedded in paraffin. Sections from the paraffin blocks were cut in 5 μm in thickness by using rotatory microtome. Then, the sections were stained with Meyer’s Hematoxylin and Eosin (H&E). The sections were protected by a thin cover slip attached to the slide with a mounting medium ‘DPX’ [<a href=\"#r-13\">13</a>]. The samples were studied with the aid of light microscope.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Photomicrographs</strong><br />\r\nPhotomicrographs were taken using ZEISS Axiocam ERc5s microscope (Germany) camera. All images were transferred to Adobe Photoshop Elements 15 (Adobe Systems, CA, USA). Only brightness and contrast were adjusted and no other adjustments were made.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Data analysis</strong><br />\r\nA statistical software package (SPSS, version 20) was used for data analysis. The descriptive data is given as mean ± standard deviation (SD). Chi-squared test was used for the analytical assessment. The differences were considered statistically significant when P values were less than 0.05 and 0.01.</p>"
},
{
"section_number": 3,
"section_title": "RESULTS",
"body": "<p><strong>Behavioral changes</strong><br />\r\nMice of group A were healthy and active without any abnormal signs during the whole experimental period. Mice of group B treated with recommended dose (5 mg/kg) were apparently normal without any abnormal sign up to 7 days of intramuscular administration of gentamicin. Mice of group C (5 mg/kg for 30 days) showed fear with less appetite, roughness of the body, apathy and weakness. However, in group D (10 mg/kg for 30 days), all the mice produced irritable behavior, roughness of the hair coat, dullness, less appetite and weakness. Mortality of the animals (12% and 50%) was found in groups C and D respectively (<a href=\"#Table-1\">Table 1</a>), but, the highest concentration was found in group D treated with 10 mg/kg for 30 days.</p>\r\n\r\n<div id=\"Table-1\">\r\n<p><strong><a href=\"https://jabet.bsmiab.org/table/178-1524741069-table1/\">Table 1</a> Table 1. </strong>Behavioral effects in the control and gentamicin treated mice.</p>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Hematological and biochemical changes</strong><br />\r\nHematologically, in group A (Control), the mean value of TEC, TLC and Hb % was 891.80±1.304 ml/cubicmeter, 8.14±0.018 thousand/cubicmeter and 8.99±0.013respectively (<a href=\"#figure1\">Figure 1</a>). The value of TEC, TLC and Hb% were decreased significantly (P<0.01) in group C (5 mg/kg for 30 days) and group D (10 mg/kg for 30 days) compared to control group (<a href=\"#figure1\">Figure 1</a>A, B, C). Biochemically, in group A (Control), the mean value of ALT was 17.28±0.130 U/L. This value was increased significantly (P<0.01) in group D (10 mg/kg for 30 days) (<a href=\"#figure1\">Figure 1</a>D).</p>\r\n\r\n<div id=\"figure1\">\r\n<figure class=\"image\"><img alt=\"\" height=\"457\" src=\"/media/article_images/2024/42/09/Figure1.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 1.</strong> Blood chemical parameters in the control and gentamicin treated mice. TEC (A), TLC (B) and Hb (C) were significantly lower in group C and D, whereas, ALT (D) was significantly higher in group C and D than that of group A and B. (** p< 0.01).</figcaption>\r\n</figure>\r\n</div>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Gross architectural changes of Liver</strong><br />\r\nReddish color with normal liver was found in Group A (control). In group B (5 mg/kg for 7 days) liver was normal after the treatment period. Whereas, congestion, dark coloration and hepatomegaly was found in group C and group D (data not shown).<br />\r\nThe mean weight of liver of control group was 2.00±0.010 g. Whereas, in group C and group D these parameters were 2.03±0.001** and 2.54±0.039** g respectively. The weight of liver was increased significantly (p<0.01) in group C (5 mg/kg for 30 days) and more significantly (p<0.01) in group D (10 mg/kg for 30 days) (<a href=\"#figure2\">Figure 2</a>).</p>\r\n\r\n<div id=\"figure2\">\r\n<figure class=\"image\"><img alt=\"\" height=\"190\" src=\"/media/article_images/2024/42/09/Figure2.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 2.</strong> Weight and size of the liver in the control and gentamicin treated mice. Both Weight (A) and size (B) of the liver were significantly increased in group C and D than that of group A and B. (** p< 0.01).</figcaption>\r\n</figure>\r\n</div>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Microscopic architectural changes of Liver</strong><br />\r\nIn liver of group A (control) and group B (pharmacological dose 7 days treatment) liver parenchyma appeared as centrally located central vein and many hepatocytes surrounding the central vein many hepatocytes (<a href=\"#figure3\">Figure 3A, 3B</a>). Long term administration of Gentamicin with pharmacological dose (group C) induced marked congestion around central vein and lymphocytic infiltration around bile canaliculi. (<a href=\"#figure3\">Figure 3C, 3D</a>). In addition to these alterations, in case of higher dose with chronic treatment (group D), dilatation of sinusoids, appearance of inclusion bodies in hepatic parenchyma, tissue destruction and irregularity of the size of hepatocytes were also found in the liver (<a href=\"#figure3\">Figure 3E, 3F, 3G</a>).</p>\r\n\r\n<div id=\"figure3\">\r\n<figure class=\"image\"><img alt=\"\" height=\"447\" src=\"/media/article_images/2024/42/09/Figure3.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 3. </strong>Photomicrograph of H & E stained histological section of liver of the (A) Control, Group A, or gentamicin injected mice with different dose rate (B) Group B, (C-D) Group C and (E-G) Group D.<br />\r\n(A) showing intact liver parenchyma with centrally located central vein. (B) also showing intact liver parenchyma. (C-D) showing congestion around central vein and lymphocytic infiltration around bile canaliculi (black arrows), (E-G) showing dilatation of sinusoids (E), inclusions bodies in hepatic parenchyma (black arrow) (F) and tissue destruction and irregularity of liver hepatocytes (G). CV, central vein. Scale bar = 20 µm.</figcaption>\r\n</figure>\r\n</div>"
},
{
"section_number": 4,
"section_title": "DISCUSSION",
"body": "<p>In our present study, swiss albino mice were used to observe the morphological and blood chemical alterations of liver after Gentamicin administration.<br />\r\nGentamicin is commonly used as a therapeutic agent against infections. But long-term exposure of gentamicin in 30% cases may induce hepatorenal toxicity [<a href=\"#r-2\">2</a>, <a href=\"#r-8\">8</a>]. In case of liver, hepatocytes are hexagonal liver cell that contains many metabolic enzymes. Liver damage may exert to pour these enzymes into plasma and can be useful for the determination of liver damage. After use of Gentamicin is responsible for increased production of reactive oxygen species ROS associated with an increase in lipid peroxidation which takes place in the cell membranes or tissues. Lipid peroxidation is an oxidative stress whereas increased production of ROS decreased antioxidants which lead to an imbalance between oxidant and antioxidant status and ultimately leading to cellular damage [<a href=\"#r-8\">8</a>, <a href=\"#r-14\">14</a>].<br />\r\nIntramuscular administration of Gentamicin in higher doses showed, roughness of the body, apathy, loss of appetite and weakness. Other groups were observed similar findings when 10 dogs received Gentamicin 10 mg/ kg IM 3 times a day for 14 days [<a href=\"#r-14\">15</a>; <a href=\"#r-16\">16</a>]. However, they also found diarrhea and vomition following administration of Gentamicin in dog. In our present study, highest concentration of mortality was found in group D (10mg/kg for 30 days). Whereas death of new born in rabbit was also reported following low dose (20mg/kg) of intramuscular injection of Gentamicin during gestation period [<a href=\"#r-17\">17</a>].<br />\r\nVarious blood chemical parameters were performed for evaluation of the functions of the organ such as TEC, TLC, Hb% and ALT. In the present study, a significant increase of Alanine amino transferasewas observed and the increased ALT is essential indicator of initial hepatocellular damage [<a href=\"#r-18\">18</a>]. It was postulated that Gentamicin treatment caused elevation in serum urea, creatinine concentration and ALT (Alanine amino transferase) activity associated with pathological changes in liver and kidney [<a href=\"#r-19\">19</a>]. Pathological lesions in the organs and chemical changes in the blood were more severe in diabetic gentamicin treated rats. It was showed that gentamicin could induce renal toxicity and significant increase in the level of ALT [<a href=\"#r-20\">20</a>]. The recorded increased level of ALT indicates functional disorders of the liver as postulated by another researcher [<a href=\"#r-21\">21</a>]. Increased level of ALT due to Gentamicin treatment induced oxidative injury causing tissue damage. This finding is in accord with that of [<a href=\"#r-22\">22</a>] who also reported similar results. In the present study, intramuscular administration of Gentamicin in 3 different doses (5mg/kg for 7 days, 5mg/kg for 30 days and 10 mg/kg for 30 days) significantly reduced the TEC, TLC and Hb%. Similar findings were reported by [<a href=\"#r-23\">23</a>] that long term exposure of Gentamicin in high dose affects the haemopoietic cells in the bone marrow and decrease erythrocyte production. From this study, congestion, dark coloration and hepatomegaly was found in treated group (5 mg/kg for 30 days and 10 mg/kg for 30 days). Gentamicin induced hepatomegaly with decrease the hepatic function in treated animal after intramuscular administration of 75 mg gentamicin /kg for 15 days in rabbits [<a href=\"#r-24\">24</a>].<br />\r\nHistomicrograph study reveals that mice treated with gentamicin with high doses for long term were showed congestion in central vein, lymphocytic infiltration in liver parenchyma and destruction of tissue architecture particularly dilatation of sinusoids. The tissue changes seen in the present work confirmed with the findings of previous work [<a href=\"#r-19\">19</a>; <a href=\"#r-25\">25</a>]. The cellular organization of mouse liver was studied using light and electron microscopy. Gentamicin treated mouse showed that approximately 35% of the hepatocytes contained two nuclei; none of the Kupffer or Ito cells were double nucleated. The presence of canaliculi and a bile duct system appear similar to that reported for other mammalian species [<a href=\"#r-26\">26</a>; <a href=\"#r-27\">27</a>].<br />\r\nIn conclusion, our present study clarified that long-term treatment of gentamicin with either pharmacological or high dose in swiss albino mice showed a fair degree of reduced food intake, increased mortality, induced significant increase of alanine aminotransferase and caused derangement of liver function with concomitant changes in the histological structures of that organ. The present study suggests that we should have conscious about taking of antibiotic in major or minor issues. The present study may be considered as an experimental base of the relevant human studies.</p>"
},
{
"section_number": 5,
"section_title": "ACKNOWLEDGEMENT",
"body": "<p>The authors extend their appreciation to the Ministry of Science and Technology, Bangladesh (MoST; Project no. BS 228/2015-16) for funding the research works.</p>"
},
{
"section_number": 6,
"section_title": "AUTHOR CONTRIBUTIONS",
"body": "<p>NJ and MRI designed the experiment. NJ, TA and NS performed the experiments; NJ, MRJ and MRI analyzed the data; NJ wrote the draft, MRJ and MRI critically revised the manuscript; MRI supervised the study.</p>"
},
{
"section_number": 7,
"section_title": "CONFLICT OF INTERESTS",
"body": "<p>The authors declare no conflict of interest.</p>"
}
],
"figures": [
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/42/09/Figure1.jpg",
"caption": "Figure 1: Blood chemical parameters in the control and gentamicin treated mice. TEC (A), TLC (B) and Hb (C) were significantly lower in group C and D, whereas, ALT (D) was significantly higher in group C and D than that of group A and B. (** p 0.01).",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/42/09/Figure2.jpg",
"caption": "Figure 2: Weight and size of the liver in the control and gentamicin treated mice. Both Weight (A) and size (B) of the liver were significantly increased in group C and D than that of group A and B. (** p 0.01).",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/42/09/Figure3.jpg",
"caption": "Figure 3: Photomicrograph of H & E stained histological section of liver of the (A) Control, Group A, or gentamicin injected mice with different dose rate (B) Group B, (C-D) Group C and (E-G) Group D.\r\n(A) showing intact liver parenchyma with centrally located central vein. (B) also showing intact liver parenchyma. (C-D) showing congestion around central vein and lymphocytic infiltration around bile canaliculi (black arrows), (E-G) showing dilatation of sinusoids (E), inclusions bodies in hepatic parenchyma (black arrow) (F) and tissue destruction and irregularity of liver hepatocytes (G). CV, central vein. Scale bar = 20 µm.",
"featured": false
}
],
"authors": [
{
"id": 32,
"affiliation": [
{
"affiliation": "Department of Anatomy & Histology, Faculty of Veterinary Medicine and Animal Science, Bangabandhu Sheikh Mujibur Rahman \r\nAgricultural University, Gazipur-1706, Bangladesh"
}
],
"first_name": "Nure",
"family_name": "Jannat",
"email": null,
"author_order": 1,
"ORCID": "https://www.researchgate.net/profile/Nure-Jannat-3",
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 20
},
{
"id": 33,
"affiliation": [
{
"affiliation": "Department of Anatomy and Histology, Faculty of Veterinary Science, Bangladesh Agricultural University, Mymensingh 2202, Bangladesh"
}
],
"first_name": "Tanjina",
"family_name": "Amin",
"email": null,
"author_order": 2,
"ORCID": "https://orcid.org/0000-0001-9699-3374",
"corresponding": false,
"co_first_author": false,
"co_author": true,
"corresponding_author_info": "",
"article": 20
},
{
"id": 34,
"affiliation": [
{
"affiliation": "Department of Anatomy and Histology, Faculty of Veterinary Science, Bangladesh Agricultural University, Mymensingh 2202, Bangladesh"
}
],
"first_name": "Nasrin",
"family_name": "Sultana",
"email": null,
"author_order": 3,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": true,
"corresponding_author_info": "",
"article": 20
},
{
"id": 35,
"affiliation": [
{
"affiliation": "Department of Anatomy and Histology, Faculty of Veterinary Science, Bangladesh Agricultural University, Mymensingh 2202, Bangladesh"
}
],
"first_name": "Mir Rubayet",
"family_name": "Jahan",
"email": null,
"author_order": 4,
"ORCID": "https://scholar.google.co.jp/citations?user=04BYdR4AAAAJ&hl=en",
"corresponding": false,
"co_first_author": false,
"co_author": true,
"corresponding_author_info": "",
"article": 20
},
{
"id": 36,
"affiliation": [
{
"affiliation": "Department of Anatomy and Histology, Faculty of Veterinary Science, Bangladesh Agricultural University, Mymensingh 2202, Bangladesh"
}
],
"first_name": "M Rafiqul",
"family_name": "Islam",
"email": "rafiqah77@yahoo.com",
"author_order": 5,
"ORCID": null,
"corresponding": true,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "M Rafiqul Islam, DVM, MS, PhD, Professor, Department of Anatomy and Histology, Faculty of Veterinary Science,\r\nBangladesh Agricultural University, Mymensingh 2202, Bangladesh, Email: rafiqah77@yahoo.com",
"article": 20
}
],
"views": 1589,
"downloads": 153,
"references": [
{
"id": 190,
"serial_number": 1,
"pmc": null,
"reference": "Ayatollahi J. Evaluation of knowledge and activities of medical students in the last two yearsof their education about chemoprophylaxis following contact with infectious diseases.Int J Clin Implant Dent. 2005;9 (26): 54-9.",
"DOI": null,
"article": 20
},
{
"id": 191,
"serial_number": 2,
"pmc": null,
"reference": "Stojiljkovic N, Stoiljkovic M. Micromorphological characteristics of the liver andbiochemical analyses in the blood of rats treated by gentamicin and verapamil. Acta MedMedianae. 2006;45 (2): 5-9.",
"DOI": null,
"article": 20
},
{
"id": 192,
"serial_number": 3,
"pmc": null,
"reference": "Gilbert DN, Mandell GL, Bennett & Dolin R. Aminoglycosides in principles and practice of infectious diseases. 5th edition. New York. 2000, pp 307-36.",
"DOI": null,
"article": 20
},
{
"id": 193,
"serial_number": 4,
"pmc": null,
"reference": "Craig W. Pharmacokinetic/pharmacodynamic parameters; rational for antibacterial dosing of mice and men. Clinical Infectious Diseases. 1998, pp 261-10.",
"DOI": null,
"article": 20
},
{
"id": 194,
"serial_number": 5,
"pmc": null,
"reference": "Hossain MM, Glass RI, Khan MR. Antibiotic use in a rural community in Bangladesh.Int J of Epidemiol.1982;11:402-5.",
"DOI": null,
"article": 20
},
{
"id": 195,
"serial_number": 6,
"pmc": null,
"reference": "Gonzelman GM. Pharmacotherapeutics of aminoglycosides antibiotics. Am JRenal Med. 1980; 5:1076-1078.",
"DOI": null,
"article": 20
},
{
"id": 196,
"serial_number": 7,
"pmc": null,
"reference": "Rieviere JE, Coppoc GL. Determination of cerebrospinal fluid Gentamicin in the beagleusing an indwelling cerebral ventricular cannula. Chemotherapy.1981;27:309-312.",
"DOI": null,
"article": 20
},
{
"id": 197,
"serial_number": 8,
"pmc": null,
"reference": "Masakazu K, Yoshiko E, Masashi E. Acquired resistance of Listeria monocytogenes in andescaped from liver parenchymal cells to gentamicin is caused by being coated with their plasma membrane. Microb Infect. 2014; 16 (3):237-243.",
"DOI": null,
"article": 20
},
{
"id": 198,
"serial_number": 9,
"pmc": null,
"reference": "Zimmerman HJ. Aminoglycosides. Hepatic injury from the treatment of infectious and parasitic diseases. In, Zimmerman HJ. Hepatotoxicity: the adverse effects of drugs and other chemicals on the liver, 2nd ed. Philadelphia, Lippincott. 1999, pp 589.",
"DOI": null,
"article": 20
},
{
"id": 199,
"serial_number": 10,
"pmc": null,
"reference": "Wojciech L, Vincent LP. Ternary Complexes of Gentamicin with Iron and Lipid CatalyzeFormation of Reactive Oxygen Species. Chem Res Toxicol. 2005;18(12):357- 364.",
"DOI": null,
"article": 20
},
{
"id": 200,
"serial_number": 11,
"pmc": null,
"reference": "Yang C, Du X, Han Y. Renal cortical mitochondria are the source of oxygen free radicalsenhanced by gentamicin. Ren Fail. 1995; 17:21-26.",
"DOI": null,
"article": 20
},
{
"id": 201,
"serial_number": 12,
"pmc": null,
"reference": "May Y H, Jochen S. Formation of a cytotoxic metabolite from gentamicin by liver.Biochem Pharmacol.1990; 40(11): 11-14.",
"DOI": null,
"article": 20
},
{
"id": 202,
"serial_number": 13,
"pmc": null,
"reference": "Luna L G. Manuals of histologic staining methods of the armed forces institute ofpathology, 3rd edition, McGraw Hill Book Company, New York. 1968.",
"DOI": null,
"article": 20
},
{
"id": 203,
"serial_number": 14,
"pmc": null,
"reference": "Nayma S, Sadia CSM, Tanveer HP, Jesmine A. Effects of Ashwagandha (Withaniasomnifera) root extract on some serum liver marker enzymes (AST, ALT) In Gentamicinintoxicated rats. J Bangladesh Soc Physiol. 2012; 7(1):1-7.",
"DOI": null,
"article": 20
},
{
"id": 204,
"serial_number": 15,
"pmc": null,
"reference": "Dantas AFM, Kommers GD, Hennemann CRA. Experimental gentamicin toxicosis in dogs. Cienc Rural. 1997; 27(3): 451-456.",
"DOI": null,
"article": 20
},
{
"id": 205,
"serial_number": 16,
"pmc": null,
"reference": "Aguiar HCR, Silva CF, Schoenau W, Kommers GD, Silva PA, Leitzka MRM, DE-AguirHCR, De-Silva PA. Urinary gammaglutamyltranspeptidase activity,urinalysis, BUN and Creatinine serum dosage as an auxiliary diagnostic means in dog nephrotoxicity induced byaminoglycosides. Ciencia- Rural. 1997; 27(2): 237-244.",
"DOI": null,
"article": 20
},
{
"id": 206,
"serial_number": 17,
"pmc": null,
"reference": "Lichthorn M. Clinical study on the safety of parentral antibiotic treatment for growing,pregnant and lactating rabbits. 1985, pp:163.",
"DOI": null,
"article": 20
},
{
"id": 207,
"serial_number": 18,
"pmc": null,
"reference": "Michalowicz J, Duda W. Phenols- sources and toxicity. Pol J Environment Stud. 2007; 16 (3):347-362.",
"DOI": null,
"article": 20
},
{
"id": 208,
"serial_number": 19,
"pmc": null,
"reference": "Atef M, Arbid MS, Hanafy MSM. Comparison of gentamicin toxicity in normal and diabetic rats. Acta vet Hung. 1992; 40(1-2):107-111.",
"DOI": null,
"article": 20
},
{
"id": 209,
"serial_number": 20,
"pmc": null,
"reference": "Kadkhodaee M, Khastar H, Faghihi M, Ghaznavi R, Zahmatkesh M. Effect of cosupplementation of vitamin E and C on gentamicin induced nephrotoxicity in rats. Exp Physiol. 2005;90(4):571-576.",
"DOI": null,
"article": 20
},
{
"id": 210,
"serial_number": 21,
"pmc": null,
"reference": "Mayne PD. Clinical Chemistry in Diagnosis and Treatment, Sixth ed. New York,USA.1994, pp:478.",
"DOI": null,
"article": 20
},
{
"id": 211,
"serial_number": 22,
"pmc": null,
"reference": "Lipsky JJ, Cheng L, Sacktor B, Leitman PS. Gentamicin uptake by renal brush bordermembrane vesicles. J. Pharmacol. Clin. Ther. 1980; 215:390-3.",
"DOI": null,
"article": 20
},
{
"id": 212,
"serial_number": 23,
"pmc": null,
"reference": "Lijana RC, Williams MC. The effects of antibiotics on hemolytic behavior of red blood cell.Cell Biophys. 1986;8(4): 223-42.",
"DOI": null,
"article": 20
},
{
"id": 213,
"serial_number": 24,
"pmc": null,
"reference": "Barza M, Ioannidis J, Cappelleri. Single or multiple daily doses of aminoglycoside forinterpretation of renal and medullary concentration. A meta-analysis. 1996, pp:338-345.",
"DOI": null,
"article": 20
},
{
"id": 214,
"serial_number": 25,
"pmc": null,
"reference": "Mahmood DH, Waters A. Comparative study of uranyl nitrate and cisplatin induced renalfailure in rat. European Journal of Drug Metabolism and Pharmacology.1994; 91: 327-336.",
"DOI": null,
"article": 20
},
{
"id": 215,
"serial_number": 26,
"pmc": null,
"reference": "Blouin A, Bolender RP, Weibel ER. Distribution of organelles and membranes betweenhepatocytes and nonhepatocytes in the rat liver parenchyma; A stereological study. J Cell Biol.1977; 72:441–455.",
"DOI": null,
"article": 20
},
{
"id": 216,
"serial_number": 27,
"pmc": null,
"reference": "Bouwens L, Baekeland M, DeZanger R, Wisse E. Quantitation, tissue distribution andproliferation kinetics of Kupffer cells in normal liver.Hepatology.1986;6:718–722.Hepatology.1986;6:718–722.",
"DOI": null,
"article": 20
}
]
},
{
"id": 24,
"slug": "178-1520836022-morphometry-and-expression-of-immunoglobulins-containing-plasma-cells-in-the-harderian-gland-of-birds",
"featured": false,
"slider": false,
"issue": "Vol1 Issue2",
"type": "review_article",
"manuscript_id": "178-1520836022",
"recieved": "2018-03-05",
"revised": null,
"accepted": "2018-04-11",
"published": "2018-05-05",
"pdf_file": "https://jabet.bsmiab.org/media/pdf_file/2023/14/178-1520836022.pdf",
"title": "Morphometry and expression of immunoglobulins-containing plasma cells in the Harderian gland of Birds",
"abstract": "<p>Johann Jacob Harder first described the Harderian gland in 1694 in deer. It is found in most terrestrial animals and is located within the variable aspects of the orbit. It is believed that this gland is involved in diverse functions. Among these, it has been held to be a site of immune response, a source of thermoregulatory lipids and pheromones, act as photoprotective organ as well as part of a retinal-pineal axis. In birds, this glad was reported first in sparrow in 1918. The Harderian gland is covered by capsule and the connective tissue septa that divide the gland into numerous unequal-sized lobes and lobules. Plasma cells are found in the interacinar space and the lumina of lobules. The recent studies suggest that the Harderian gland act as an immunopotent organ in birds, and that the gland in scavenging birds contains more immunoglobulin-containing plasma cells due to their scavenging nature. Moreover, this gland shows considerable species/strain differences in terms of macro anatomy, microanatomy as well as in the dynamics of immunoglobulin-containing plasma cells among different birds. In this review, these species and strain differences are discussed based on recent studies and several goals of future research are identified.</p>\r\n<p> </p>",
"journal_reference": "J Adv Biotechnol Exp Ther. 2018; 1(2) : 55-60",
"academic_editor": "Dr. Md Mahmodul Hasan Sohel, Erciyes University, Turkey.",
"cite_info": "Jahan MR, Islam MN, Khan MZI, etal. Morphometry and expression of immunoglobulins-containing plasma cells in the Harderian gland of Birds. J Adv Biotechnol Exp Ther. 2018; 1(2) : 55-60.",
"keywords": [
"birds",
"Harderian gland",
"immunohistochemistry",
"species differences",
"anatomy"
],
"DOI": "10.5455/jabet.2018.d10",
"sections": [
{
"section_number": 1,
"section_title": "INTRODUCTION",
"body": "<p>Birds are continuously exposed to a wide spectrum of potential environmental immunomodulators including physical and chemical factors as well as various microorganisms and antigenic particles, whether occurring naturally or deliberately introduced. In addition, temperature, housing, air quality, diet, environment contaminants, feed additives, therapeutics and vaccines are significant categories of immunomodulators. To cope with the diversity of potentially harmful agents on one hand, and to mount a protective immune response on the other, birds possess an array of humoral and cell mediated immune mechanisms that destroy infected cells or pathogens and mount an adaptive immune response. For this, immunocompetent defense system is the decisive prerequisite [<a href=\"#r-1\">1</a>, <a href=\"#r-2\">2</a>]. The lymphoid tissue of the chicken is divided into “central” and ” peripheral” ones. The central lymphoid tissue includes bursa of Fabricius and thymus. The peripheral lymphoid tissue includes the spleen, cecal tonsils and all the mucosa-associated lymphoid tissues including respiratory tract, genitourinary tract, alimentary tract and head associated lymphoid tissues that consists of Harderian gland [<a href=\"#r-2\">2</a>, <a href=\"#r-3\">3</a>, <a href=\"#r-4\">4</a>, <a href=\"#r-5\">5</a>, <a href=\"#r-6\">6</a>, <a href=\"#r-7\">7</a>]<br />\r\nThe Harderian gland was first described by Johann Jacob Harder in deer on 1694 and is found in most of the terrestrial vertebrates, amphibians, reptiles, birds and mammals [<a href=\"#r-8\">8</a>]. The Harderian gland forms a unitary structure, which is firmly attached to the medial part of the orbit, and the duct of this gland opens usually on the surface of the nictitating membrane, which is a characteristic feature of this gland and distinguishes from lacrimal gland and the other ocular glands. The later ones are normally classified as a cluster of glandular tissue actually within the nictitating membrane, whereas, Harderian gland is often surprisingly large, in some cases larger that eye itself [<a href=\"#r-9\">9</a>], with diverse functions (<a href=\"#figure1\">Figure 1</a>).<br />\r\nAvian Harderian gland was first described in sparrow (Passer domesticus) in 1918, which was located at the variable aspects of orbit [<a href=\"#r-3\">3</a>, <a href=\"#r-9\">9</a>, <a href=\"#r-10\">10</a>]. The Harderian gland is relatively larger in the fowl, much larger than the lacrimal gland. In birds, the usual function of this gland is to lubricate the surface of the eyeball and nictitating membrane [<a href=\"#r-11\">11</a>]. In addition, the Harderian gland possesses numerous large plasma cells specific for anti-immunoglobulin (Ig) A, -IgG or -IgM marker in the interstitial stroma of the chicken that plays a part in immunological defense of the para ocular region [<a href=\"#r-10\">10</a>, <a href=\"#r-12\">12</a>, <a href=\"#r-13\">13</a>]. Many other authors also suggest that the Harderian gland is responsible for the local immunity of the eye orbit [<a href=\"#r-14\">14</a>, <a href=\"#r-15\">15</a>]. At the same time, it is accepted that the gland is a peripheral lymphoepithelial organ, which together with the spleen, the bursa of Fabricius and the caecal tonsils form a system of avian organs that determines both the general and the local immunity [<a href=\"#r-16\">16</a>, <a href=\"#r-17\">17</a>, <a href=\"#r-18\">18</a>]. The major classes of Igs produced in chicken Harderian gland are probably related to the secretory feature of this gland [<a href=\"#r-19\">19</a>, <a href=\"#r-20\">20</a>]<br />\r\nTill to date, Harderian gland of birds has been studied in different species or strains of birds such as chicken, duck, goose, ostrich and Guinea Fowl. The present review is focused on the species or strain differences on macro anatomical, micro anatomical and immunohistochemical studies particularly the differences in the frequencies and distribution of Ig-containing plasma cells.</p>\r\n\r\n<div id=\"figure1\">\r\n<figure class=\"image\"><img alt=\"\" height=\"448\" src=\"/media/article_images/2024/36/08/178-1520836022-Figure1.jpg\" width=\"564\" />\r\n<figcaption><strong>Figure 1. </strong>Schematic diagram for Harderian gland and its possible diverse function. This figure is adapted from [<a href=\"#r-3\">3</a>], [<a href=\"#r-9\">9</a>] and [<a href=\"#r-10\">10</a>].</figcaption>\r\n</figure>\r\n</div>"
},
{
"section_number": 2,
"section_title": "MACROANATOMY OF HARDERIAN GLAND",
"body": "<p><strong>Location</strong><br />\r\nHarderian gland of bird shows considerable species differences in location. In White Leghorn chickens the Harderian gland is located in the ventro-medail aspect of the eyeball extending rostrally from the optic nerve [<a href=\"#r-23\">23</a>].In contrast, it has been reported that Harderian gland of native chickens of Bangladeshis situated on the dorsal posterior surface of the eyeball occupying the considerable part of the orbit [<a href=\"#r-3\">3</a>, <a href=\"#r-4\">4</a>, <a href=\"#r-10\">10</a>]. While in Rook, this gland is located in the ventral and posterior medial to the eyeball [<a href=\"#r-3\">3</a>]. In the Canadian ostrich, the Harderiangland is located ventromedially around the posterior part of the eyeball [<a href=\"#r-24\">24</a>]. In duck, this gland is located in the anteriomedial part of the orbit [<a href=\"#r-25\">25</a>]</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Shape</strong><br />\r\nIn White leghorns, the harderian gland is hourglass shaped and in Rook it is tongue shaped [<a href=\"#r-3\">3</a>]. In native chickens of Bangladesh, this gland is triangular in shape [<a href=\"#r-3\">3</a>, <a href=\"#r-4\">4</a>]. Whereas, in Canadian ostrich, the Harderian gland is flattened, oval shaped, irregular in outline and pointed in the dorsal end [<a href=\"#r-24\">24</a>].</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Size</strong><br />\r\nThe size of the Harderian gland is also varied among the different types of the birds. In native chickens, the Harderian gland of male bird is 9.2 mm in length and 5.1 mm in breadth, whereas, the Harderian gland of female bird is 9.0 mm in length and 3.2 mm in breadth [<a href=\"#r-4\">4</a>]. The mean dimension of the Harderian gland in White leghorn is 17.66 mm in length and 6.2 mm in breadth [<a href=\"#r-23\">23</a>], which is almost double than that of native chickens. Whereas, Harderian gland of rook is bigger than that of White leghorn [<a href=\"#r-3\">3</a>].It is stated that the length and breadth of Harderian gland in rook is 18mm and 8.6 mm respectively. Interestingly, the size of the Harderian gland in Canadian ostrich is much bigger and it is 35.30 mm in length and 15.65 mm in breadth [<a href=\"#r-24\">24</a>]. There are no significant differences in size of the left and right gland, which is similar in the report of native chickens of Bangladesh [<a href=\"#r-4\">4</a>].</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Color</strong><br />\r\nThe Harderian gland is covered with a thin layer of fat and after removing the fat this gland of native chickens shows brownish in color [<a href=\"#r-4\">4</a>], while in broiler it is reddish in color [<a href=\"#r-3\">3</a>]. The color of Harderian gland of White leghorn bird is pale pink and rook it is pink yellow [<a href=\"#r-23\">23</a>] and in Canadian ostrich, it is light pink in color [<a href=\"#r-24\">24</a>].</p>"
},
{
"section_number": 3,
"section_title": "MICROANATOMY OF HARDERIAN GLAND",
"body": "<p>The Harderian gland of birds is surrounded with a thick connective tissue capsule. From the capsule, the trabeculae enter into the gland, dividing the gland into different sized lobes and lobules. These inter-lobular trabeculae are thinner compared to the capsule surrounded the whole gland and contain collagen fibers, fibroblasts, blood vessels and nerve fibers. The smallest secretory unit of the Harderian gland is called acini lined by simple columnar epithelial cells [<a href=\"#r-3\">3</a>, <a href=\"#r-10\">10</a>, <a href=\"#r-11\">11</a>,<a href=\"#r-24\"> 24</a>, <a href=\"#r-25\">25</a>]. There is a variation in length of the lobules in between broiler male and female, and in between broiler male and native male, however, their breadth does not vary [<a href=\"#r-3\">3</a>]. The lumen of the lobules is irregular in native chicken and regular in broiler. This variation in size of the histological lobule of Harderian gland of chickens might be due to strain differences. The lumen of the acini in broiler is spherical and regular or elongated, and the cell boundaries were distinctly visible. Whereas, in native male, the acini are lined by tall single columnar epithelium and the lumen of the acini in these strains of chickens were elongated, irregular and narrower, and the cell boundaries are not distinctly visible. This luminal variation, in regards to its cellular contents and shape in between broiler and native chicken is also possibly due to strain differences [<a href=\"#r-3\">3</a>, <a href=\"#r-10\">10</a>].</p>"
},
{
"section_number": 4,
"section_title": "LYMPHOID CELLS IN THE HARDERIAN GLAND",
"body": "<p>Different types of lymphoid cells such as macrophages, lymphocytes, plasma cells as well as granulocytes are found in the subepithelial layer of the Harderian gland of birds [<a href=\"#r-26\">26</a>, <a href=\"#r-27\">27</a>]. Although, these cells are often densely aggregated in the central regions of the lobules, individual cells are also occurred near the periphery of the Harderian gland. It has been reported that pre-plasma cells form a desmosome-like contact with macrophages and junctional complexes with each other. In addition, the lymphoid cells were sometimes embedding in a dense homogeneous material, which is also found in the intercellular spaces of the secretory epithelium [<a href=\"#r-28\">28</a>]. In contrast, other research group described that the lymphoid substance of the chicken’s Harderian gland organized in two histologically distinct compartments [<a href=\"#r-29\">29</a>]. Light and electron microscopical investigations reveal that the lymphoid structure of the Harderian gland of chickens organizes in different histological frameworks. In the head the surface epithelium of the central canal can be classified as a lymphoepithelial tissue, which covers the dense lymphoid substance. It consists of small and medium-sized lymphocytes, dendritic-like cells, and occasional macrophages. High endothelial venules are associated with intense lymphocyte migration and homing that give circumstantial evidence for a T-dependent region, as found in a secondary lymphoid organ. The B-dependent germinal centers are also common structural units of the head region’s lymphoid substance. The body of the gland is loaded with plasma cells of different maturation stages. They immigrate into the epithelium of the central canal and produce IgM and IgA and IgG [<a href=\"#r-28\">28</a>, <a href=\"#r-30\">30</a>]. This plasmocytic region is accounted for the immunosurveillance on the conjunctiva and in the upper respiratory tract through antibody production against bacterial or parasitic infections. In both the head and body regions of the gland, anti-B-L (anti-Ia) antibody recognizes scattered elongated cells that might represent dendritic cells. The immunological relationship between the two histologically different parts of the Harderian gland is unknown, but it is speculated that the dense lymphoid tissue with high endothelial venule receive the blood-borne, immunologically mature, but uncommitted B cells. By the influence of local antigen stimulus, these B cells transform to plasma cells, which gradually appear in the body of the gland. The lymphoid structures of the head and the body fulfill the function of secondary and tertiary lymphoid organs, respectively [<a href=\"#r-29\">29</a>]. The dynamics of IgA-, IgG- and IgM-containing plasma cells also shows strain variations that discussed below.</p>"
},
{
"section_number": 5,
"section_title": "IMMUNOGLOBULINS-CONTAINING PLASMA CELLS IN THE HARDERIAN GLAND",
"body": "<p>The immunoglobulins are secreted by Igs- containing plasma cells that perform local or systemic defenses in the body. It has been shown that in the native chickens the frequency of the population of IgA-positive cells is higher followed by IgG and IgM-positive cells and in the broiler chickens the frequency of IgM-positive cells is more followed by IgA- and IgG-positive cells [<a href=\"#r-10\">10</a>]. These observations are similar to the study made by different research groups in chickens [<a href=\"#r-31\">31</a>, <a href=\"#r-32\">32</a>] but varied greatly with the observations in the White Leghorn chickens [<a href=\"#r-12\">12</a>]. It has been revealed thatin White Leghorn chicken IgG- is more than IgA- and IgM-positive cells. In addition, the frequency of the population of Ig- containing plasma cells is varied between the sexes in the same strain, and also in between different strain of chickens [<a href=\"#r-10\">10</a>]. In the broiler, IgA- and IgG- positive cells are more in the male, whereas, IgM-containing cells are more in female.In the native chickens the frequency of IgA- and IgM-containing cells are more in the female, in contrast, the IgG –positive cells are higher in the male. The frequencies of Ig-containing plasma cells are also varied between sexes. The population of IgA- and IgG-containing cells is significantly higher in the native male than the broiler male chickens. In addition, the frequency of IgA- and IgG-containing cells is more in native female than the broiler female. It has been speculated that these variations concerning the frequency of occurrences of the plasma cells positive for the different classes of immunoglobulins in the Harderian gland of birds might be related to age, strain, species, health condition and their the condition of their surrounding environment. Interestingly, significant differences in lymphatic cells components are also observed between the Harderian gland of wild pheasants and hybrid pheasants [<a href=\"#r-33\">33</a>]. It has been reported that the common wild pheasant presented a higher number of immunoglobulin-producing plasma cells. The hybrid pheasants have a lower number of plasma cells, but a higher number of lymphocytes in the gland tissue in comparison with the common pheasant [<a href=\"#r-33\">33</a>]. In addition, in domestic duck, the IgM-containing plasma cells are the most abundant than other classes of immunoglobulins into the epithelium of the Harderian gland. In contrast, IgY- or IgA-positive cells are predominant in the lamina propria [<a href=\"#r-25\">25</a>].</p>"
},
{
"section_number": 6,
"section_title": "SECRETORY MECHAMISM OF PLASMA CELL IN THE HARDERIAN GLAND",
"body": "<p>Interestingly enough, strain variations are also observed in the mode of release of Ig-containing plasma cells. It is reported that secretory IgA-, IgG-, and IgM-containing plasma cells are released by the disintegration of the acinar cells at the apical part of the lobules of the broiler but by dissolution or holocrine mode of secretion in the native chicken [<a href=\"#r-10\">10</a>]. The secretory mechanism is merocrine in rabbit [<a href=\"#r-33\">33</a>] merocrine to apocrine in the other avian species [<a href=\"#r-25\">25</a>]. In a very recent study in Canadian ostrich it is also observed apocrine in nature [<a href=\"#r-24\">24</a>]. Among the available studies on native chickens of Bangladesh showed holocrine type of secretion for plasma cell in the Harderian gland.</p>"
},
{
"section_number": 7,
"section_title": "DIRECTIONS FOR FUTURE STUDY",
"body": "<p>The comparative ontogeny of Harderian gland of birds has not studied yet. The comparative trajectory of different classes of Ig-containing plasma cells in the Harderian gland of different species of birds will also need to be clarified by future study. In addition, the further knowledge, component and function of Harderian gland as a part of immune system will also need to be clarified.</p>"
},
{
"section_number": 8,
"section_title": "CONCLUSION",
"body": "<p>Harderian gland of birds, even though it is not a primary lymphoid organ as a whole, but act as an immunopotent organ, and that showed considerable species or strain variations in anatomical or histological structures as well as in the dynamics of Ig-containing plasma cells (<a href=\"#Table-1\">Table 1</a>). The wild type or domestic birds contained more Igs-containing plasma cells possibly due to their scavenging in nature. The current review could also serve as a caution that data on morphology or frequencies of Igs-containing plasma cells of Harderian gland from one species should not always be directly applied to others among birds.</p>\r\n\r\n<div id=\"Table-1\">\r\n<p><strong><a href=\"https://jabet.bsmiab.org/table/178-1520836022-Table1/\">Table-1</a> Table 1. </strong>Species differences in gross and histochemical parameters of Harderian gland of birds <sup>[3,</sup> <sup>4,</sup> <sup>9,</sup> <sup>10,</sup> <sup>11,</sup> <sup>23,</sup> <sup>2].</sup></p>\r\n</div>"
},
{
"section_number": 9,
"section_title": "CONFLICT OF INTEREST",
"body": "<p>The author declares that no conflict of interest exists.</p>"
}
],
"figures": [
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/36/08/178-1520836022-Figure1.jpg",
"caption": "Figure 1: Schematic diagram for Harderian gland and its possible diverse function. This figure is adapted from [3], [9] and [10].",
"featured": false
}
],
"authors": [
{
"id": 49,
"affiliation": [
{
"affiliation": "Department of Anatomy and Histology, Bangladesh Agricultural University, Mymensingh 2202, Bangladesh"
},
{
"affiliation": "Division of Neuroanatomy, Department of Neuroscience, Yamaguchi University Graduate School of Medicine, Ube 755-8505, Yamaguchi, \r\nJapan"
}
],
"first_name": "Mir Rubayet",
"family_name": "Jahan",
"email": "rubayet.lucky@gmail.com",
"author_order": 1,
"ORCID": "https://orcid.org/0000-0003-2867-5336",
"corresponding": true,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "Mir Rubayet Jahan, DVM, MS, PhD, Associate Professor, Department of Anatomy and Histology, Bangladesh Agricultural University, Mymensingh 2202, Bangladesh, Email: rubayet.lucky@gmail.com",
"article": 24
},
{
"id": 50,
"affiliation": [
{
"affiliation": "Division of Neuroanatomy, Department of Neuroscience, Yamaguchi University Graduate School of Medicine, Ube 755-8505, Yamaguchi, \r\nJapan"
}
],
"first_name": "Md. Nabiul",
"family_name": "Islam",
"email": null,
"author_order": 2,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 24
},
{
"id": 51,
"affiliation": [
{
"affiliation": "Department of Anatomy and Histology, Bangladesh Agricultural University, Mymensingh 2202, Bangladesh"
}
],
"first_name": "Md. Zahirul Islam",
"family_name": "Khan",
"email": null,
"author_order": 3,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 24
},
{
"id": 52,
"affiliation": [
{
"affiliation": "Division of Neuroanatomy, Department of Neuroscience, Yamaguchi University Graduate School of Medicine, Ube 755-8505, Yamaguchi, \r\nJapan"
}
],
"first_name": "Akie",
"family_name": "Yanai",
"email": null,
"author_order": 4,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 24
},
{
"id": 53,
"affiliation": [
{
"affiliation": "Division of Neuroanatomy, Department of Neuroscience, Yamaguchi University Graduate School of Medicine, Ube 755-8505, Yamaguchi, \r\nJapan"
}
],
"first_name": "Koh",
"family_name": "Shinoda",
"email": null,
"author_order": 5,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 24
}
],
"views": 1200,
"downloads": 122,
"references": [
{
"id": 67,
"serial_number": 1,
"pmc": null,
"reference": "Jeurissen SHM, Claassen E, Janse EM. Histological and functional differentiation of non-lymphoid cells in the chickens spleen. Immunology.1992; 77: 75-80.",
"DOI": null,
"article": 24
},
{
"id": 290,
"serial_number": 2,
"pmc": null,
"reference": "Jeurissen SHM, Vervelde L, Janse EM. Structure and function of lymphoid tissues of the chicken. Poultry Sci. Rev.1994; 5: 183-207.",
"DOI": null,
"article": 24
},
{
"id": 291,
"serial_number": 3,
"pmc": null,
"reference": "Jahan MR, Khan MZI, Islam MN, Akter SH.Morphometrical analysis of Harderian gland of broiler and native chickens of Bangladesh. J Bangladesh Agril Univ. 2006; 4(2): 273-278",
"DOI": null,
"article": 24
},
{
"id": 292,
"serial_number": 4,
"pmc": null,
"reference": "Jahan MR, Islam MN, Khan MZI. Gross and biometrical study of the Harderian gland in adult native chickens of Bangladesh.Int J Biol Res. 2009; 7: 31-34.",
"DOI": null,
"article": 24
},
{
"id": 293,
"serial_number": 5,
"pmc": null,
"reference": "Islam MN, Khan MZI, Jahan MR, Karim MR, Kon Y. Comparative studies of mucosa and immunoglobulin-containing plasma cells in the gastrointestinal tract of broiler and native chickens of Bangladesh. J. Poult. Sci. 2008; 45: 125-131.",
"DOI": null,
"article": 24
},
{
"id": 294,
"serial_number": 6,
"pmc": null,
"reference": "Islam MN, Khan MZI, Jahan MR, Fujinaga R, Yanai A, Kokubu K, Shinoda K. Histomorphological study on prenatal development of the lymphoid organs of native chickens of Bangladesh. Pak. Vet. J. 2012; 32: 175–178.",
"DOI": null,
"article": 24
},
{
"id": 295,
"serial_number": 7,
"pmc": null,
"reference": "Islam MN, Khan MZI, Jahan MR, Shinoda K. Developmental trajectory of the prenatal lymphoid organs in native chickens:a macro anatomical study. Asian J. Med. Biol. Res. 2017; 3(4): 432–436.",
"DOI": null,
"article": 24
},
{
"id": 296,
"serial_number": 8,
"pmc": null,
"reference": "vanGinkel FW, Tang DC, Gulley SL, Toro H. Induction of mucosal immunity in the avian Harderian gland with a replication-deficient Ad5 vector expressing avian influenza H5 hemagglutinin. Dev Comp Immunol. 2009; 33: 28-34",
"DOI": null,
"article": 24
},
{
"id": 297,
"serial_number": 9,
"pmc": null,
"reference": "Payne AP. The Harderian gland: a tercentennial review. J. Anat. 1994;185: 1-49.",
"DOI": null,
"article": 24
},
{
"id": 298,
"serial_number": 10,
"pmc": null,
"reference": "Khan MZI, Jahan MR, Islam MN, Haque Z, Islam MR, Kon Y. Immunoglobulin (Ig)-containing plasma cells in the Harderian gland in broiler and native chickens of Bangladesh. Tissue and cell. 2007; 39: 141–149.",
"DOI": null,
"article": 24
},
{
"id": 299,
"serial_number": 11,
"pmc": null,
"reference": "Mobini B. Histological and histochemical studies on the Harderian gland in native chickens. Vet Med. 2012; 8: 404-409.",
"DOI": null,
"article": 24
},
{
"id": 300,
"serial_number": 12,
"pmc": null,
"reference": "Nasrin M, Khan MZI, Siddiqi MNH, Masum MA. Mobilization of immunoglobulin (Ig)-containing plasma cells in Harderian gland, cecal tonsil and trachea of broilers vaccinated with Newcastle Diseases Vaccine. Tissue and Cell. 2013; 45: 191-197.",
"DOI": null,
"article": 24
},
{
"id": 301,
"serial_number": 13,
"pmc": null,
"reference": "Bejdic P, Avdic R. Amidzic L, Cutahija V, Hadziomerovic N, Katica A, Mlaco N. Ultrastuctural of plasma cells in harderian gland of laying hen. AntatHistolEmbryol. 2017; 47: 46-50.",
"DOI": null,
"article": 24
},
{
"id": 302,
"serial_number": 14,
"pmc": null,
"reference": "Baba T, kawata T, Masumoto K, Kajikawa T. Role of the Harderian gland in immunoglobulin A production in chicken lacrimal fluid. ResVet Sci. 1990; 49: 20-26.",
"DOI": null,
"article": 24
},
{
"id": 303,
"serial_number": 15,
"pmc": null,
"reference": "Ohshima K, Hiramatsu K. Immunohistochemical localization of three different immunoglobulin classes in the Harderian gland of young chicken. Tissue cell. 2002; 34 (2): 129-133.",
"DOI": null,
"article": 24
},
{
"id": 304,
"serial_number": 16,
"pmc": null,
"reference": "Shirama K, SatohT, Kitamura T, Yamade J.The avian Harderian gland: Morphology and Immunology. MicroscRes and Tech. 1996; 34: 16-27.",
"DOI": null,
"article": 24
},
{
"id": 305,
"serial_number": 17,
"pmc": null,
"reference": "Korbel R, Schaffer E, RavelhoferK, Kosters J. (1997). Ocular Manifestation of mycobacterium infections in bird.TierartzlParxAusg K. 1997; 25: 552-558.",
"DOI": null,
"article": 24
},
{
"id": 306,
"serial_number": 18,
"pmc": null,
"reference": "Fix AS, Arp LH. Morphologic characterization of conjunctiva- associated lymphoid tissue in chicken. Am J Vet Res. 1998; 52 (11): 1852-1859.",
"DOI": null,
"article": 24
},
{
"id": 307,
"serial_number": 19,
"pmc": null,
"reference": "Albini B, Wick G, Rose E. Immunoglobulin production in chicken Harderian gland. Int Arch Allergy.1974; 47: 23-34.",
"DOI": null,
"article": 24
},
{
"id": 308,
"serial_number": 20,
"pmc": null,
"reference": "Baba T, MasumotoK, Nishida S, kajikawa T, Mitsui. Harderian gland dependency of immunoglobulin A production in the lacrimal fluid of chicken. Immunology.1988; 65(1): 67-71.",
"DOI": null,
"article": 24
},
{
"id": 309,
"serial_number": 21,
"pmc": null,
"reference": "Rahman ML, Islam MR, Asaduzzaman M, Khan MZI. Lymphoid tissues in the digestive tract of Deshi Chicken (Gallus domesticus) in Bangladesh. Pak J Biol Sci. 2003; 6: 1145-1150.",
"DOI": null,
"article": 24
},
{
"id": 310,
"serial_number": 22,
"pmc": null,
"reference": "Islam MN, Khan MZI, Jahan MR, Fujinaga R, Shinoda K. Ontogenic development of immunoglobulins (Igs)-positive lymphocytes in the lymphoid organs of native chickens of Bangladesh. Int. J Vet Sci Med. 2013; 1: 96-101.",
"DOI": null,
"article": 24
},
{
"id": 311,
"serial_number": 23,
"pmc": null,
"reference": "Ashok P, Venkatkrishnan A, Vijayaragavan C,Ramkrishna V. Gross morphology and biometrical study of the Harderian gland in White Leghorn birds. Indian J Anim Sci. 2000; 15(2): 217-219.",
"DOI": null,
"article": 24
},
{
"id": 312,
"serial_number": 24,
"pmc": null,
"reference": "Frahmand S, Mohammadpour AA. Harderian gland in Canadian Ostrich (Struthiocamelus): A morphological and histochemical study.Antat Histol Embryol. 2015; 44: 178-185.",
"DOI": null,
"article": 24
},
{
"id": 313,
"serial_number": 25,
"pmc": null,
"reference": "Oliveria CA, Telles FL, Oliveria AG, Kalapothakis E, Goncalves-Dornelas H, Mahecha GAB. Expression of different classes of immunoglobulin in intraepithelial plasma cells of the Harderian gland of domestic ducks Anasplatyrhynchos. Vet ImmunolImmunopathol. 2006; 113: 257-266.",
"DOI": null,
"article": 24
},
{
"id": 314,
"serial_number": 26,
"pmc": null,
"reference": "Bejdic P, Avdic R. Amidzic L, Cutahija V, Tandir F, Hadziomerovic N. Developmental changes of lymphoid tissue in the Harderian gland of laying hens. Mac Vet Rev. 2014; 37: 83-88.",
"DOI": null,
"article": 24
},
{
"id": 315,
"serial_number": 27,
"pmc": null,
"reference": "Scott TR. Our current understanding of humoral immunity of poultry. Poultry Sci. 2004; 83: 574-579.",
"DOI": null,
"article": 24
},
{
"id": 316,
"serial_number": 28,
"pmc": null,
"reference": "Khan MZI, Akter SH, Islam MN, Karim MR, Islam MR, Kon Y. The Effect of Selenium and Vitamin E on the Lymphocytes and Immunoglobulincontaining Plasma cells in the Lymphoid organ and Mucosa-Associated Lymphatic Tissues of Broiler Chicken. AnatHistolEmbryol. 2008; 37: 52-59.",
"DOI": null,
"article": 24
},
{
"id": 317,
"serial_number": 29,
"pmc": null,
"reference": "Olah I, Kupper A, Kittner Z. The lymphoid substance of the chicken’s Harderian gland is organized in two histologically distinct compartments.Microsc Res Tech.1996; 34(2):166-76.",
"DOI": null,
"article": 24
},
{
"id": 318,
"serial_number": 30,
"pmc": null,
"reference": "Masum MA, Khan MZI, Nasrin M, Siddiqi MNH, Khan MZI, Islam MN, Detection of immunoglobulins containing plasma cells in the thymus, bursa of Fabricius and spleen of vaccinated broiler chickens with Newcastle disease virus vaccine, Int J Vet Sci Med. 2014; 2 (2): 103-108.",
"DOI": null,
"article": 24
},
{
"id": 319,
"serial_number": 31,
"pmc": null,
"reference": "Bienenstock J, Gauldie J. Perey DYE.Synthesis of IgG, IgA and IgM by chicken tissues.J Immunol. 1973; 111: 1112-1118.",
"DOI": null,
"article": 24
},
{
"id": 320,
"serial_number": 32,
"pmc": null,
"reference": "Klećkowska-Nawrot J, Goździewska- arłajczuk K, Kowalczyk A, Łukaszewicz E, Nowaczyk R. Structural Differences of the Harderian Gland between Common Pheasants (PhasianusColchicusTalischensis) and Hybrids of Italian Amber and Common Pheasants.Rev. Bras. Cienc. Avic. 2016; 18 (2): 309-318",
"DOI": null,
"article": 24
},
{
"id": 321,
"serial_number": 32,
"pmc": null,
"reference": "Kuhnel W. Structural and cytochemical study of Harderian gland of Druse von kaninchen, Z. Zelloforsch.Mikrosk. Amt.1971; 119: 384-404.",
"DOI": null,
"article": 24
}
]
},
{
"id": 21,
"slug": "178-1521008794-isolation-of-stem-cell-populations-from-whartons-jelly-sections-of-umbilical-cord-and-comparison-analysis-with-cord-blood-stem-cells",
"featured": false,
"slider": false,
"issue": "Vol1 Issue2",
"type": "original_article",
"manuscript_id": "178-1521008794",
"recieved": "2018-02-12",
"revised": null,
"accepted": "2018-03-12",
"published": "2018-05-05",
"pdf_file": "https://jabet.bsmiab.org/media/pdf_file/2023/06/178-1521008794.pdf",
"title": "Isolation of stem cell populations from wharton’s jelly sections of umbilical cord and comparison analysis with cord blood stem cells",
"abstract": "<p>Wharton’s jelly (WJ) is a gelatinous tissue exists in the umbilical cord contains stromal cells, termed as mesenchymal stromal cells (MSCs). Current study designed to characterize the stem cell population both within the cord blood and it’s wharton’s jelly tissue and it’s correlation to neonatal parameters. Both wharton’s jelly and cord blood and their CD34+ and CD90+ expression was enumerated by flow-cytometry. Neonatal parameters along with the mode of delivery were analyzed to correlate with CD34 and CD90 markers. From a total of 20 deliveries, CD34 and CD90 expression pattern showed varied in both samples. Gender and the expression of CD34 and CD90 markers doesn’t show any significant change. Whereas, correlation analysis of weight with CD34+ and CD90+ cell counts showed significant difference and it may have the effect on both cell counts. Neonatal parameters especially birth weight may have the effect on cell count of CD34 and CD90 in both CB and WJ. Current study provide a fast method for mesenchymal cell isolation from Wharton’s jelly tissue using the total length of the cord, which may be easily applied along with the UC blood, for the banking of UC.</p>",
"journal_reference": "J Adv Biotechnol Exp Ther. 2018; 1(2) : 36-42",
"academic_editor": "Dr. Gautam Deb, Bangladesh Livestock Research Institute, Bangladesh.",
"cite_info": "Shaik MV, K H, G S.Isolation of stem cell populations from wharton’s jelly sections of umbilical cord and comparison analysis with cord blood stem cells. J Adv Biotechnol Exp Ther. 2018; 1(2) : 36-42.",
"keywords": [
"umbilical cord",
"Wharton’s jelly",
"phenotypic characteristics",
"therapeutic applications"
],
"DOI": "10.5455/jabet.2018.d7",
"sections": [
{
"section_number": 1,
"section_title": "INTRODUCTION",
"body": "<p>In the recent years, there seems to be an unbounded interest regarding mesenchymal stem cells. MSC from the bone marrow (BM) is still considered as the gold standard for various clinical applications and stem cell engineering. But the bone marrow has limitations as MSCs source, like mesenchymal stem cell low frequency, painful procedure and the decline in MSC characteristics with donor’s age [<a href=\"#r-1\">1</a>]. Kobayashi et al., [<a href=\"#r-2\">2</a>] demonstrated that umbilical cord which may be discarded during delivery can be provided as an inexhaustible source for stem cells and they shows the similar characteristics like MSCs from bone marrow. Wharton’s jelly derived MSCs have advantageous characteristics like plenty source, rapidly available donor, non-invasive collection and painless procedure, ethically noncontroversial and doesn’t shows any teratoma [<a href=\"#r-3\">3</a>]and this may have immune privileged and immunomodulatory phenotype capable of suppressing the immune response in vitro. Whereas bone marrow MSC (bMSC) may shows notable changes with increasing patient’s age, reduction in the available cell number, longer doubling times and lower differentiation potential in vitro. Wharton’s jelly stem cells shown to be differentiated into osteoblast [<a href=\"#r-4\">4</a>], chondrocyte [<a href=\"#r-5\">5</a>], cardiomyocyte [<a href=\"#r-6\">6</a>], skeletal myoblast [<a href=\"#r-7\">7</a>], hepatocyte cells [<a href=\"#r-8\">8</a>], endothelial cell [<a href=\"#r-9\">9</a>], neural cell & adipocyte [<a href=\"#r-10\">10</a>], dopaminergic cell [<a href=\"#r-11\">11</a>] andlens fiber cell [<a href=\"#r-12\">12</a>]. Wharton’s jelly stem cells shows positive forCD44, CD105 [<a href=\"#r-7\">7</a>, <a href=\"#r-13\">13</a>], CD68 [<a href=\"#r-14\">14</a>], CD13 and CD95 markers, but negative for CD34, CD45, CD38 and CD71 markers. Wharton’s jelly MSCs are primitive MSCs than other tissue sources and theydon’t express the major histocompatibility complex (MHC) class II HLA-DR [<a href=\"#r-15\">15</a>]. The important graft-vs-host disease (GVHD) markers like CD86, CD80 and CD40 were not noticeable or weakly expressed in WJMSCs [<a href=\"#r-16\">16</a>]. CD34+ and CD90+ marker analysis was useful for the examination of the quality of cord blood for future any hematological and non-hematological disease therapeutics. Still there was little or no knowledge with regard to neonatal characteristics and their effect on CD34 and CD90 cell population in WJ and CB. Hence current study designed to analyze the cell population from complete section of umbilical cord wharton’s jelly and it’s comparison to the stem cells population derived from cord blood. Also we evaluated the cell counts of each of the marker of fresh cord blood samples and WJ sample and were correlated between their expressions with neonatal parameters.</p>"
},
{
"section_number": 2,
"section_title": "MATERIALS AND METHODS",
"body": "<p><strong>Materials</strong><br />\r\nFetal bovine serum, streptomycin, trypan blue and phosphate-buffered saline pH 7.4 (Gibco, USA). Collagenase type II, Bovine serum albumin, DMEM-F12 Medium (Sigma-Aldrich, USA).CD133, CD90 Antibodies conjugated microbeads kits and miniMACS magnetic column with separation system from Miltenyi Biotech, Germany. All chemicals & Reagents are of molecular biology grade.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Collection of Wharton’s jelly tissue material in sterile condition</strong><br />\r\nNormally, cords and it’s blood will be thrown after the delivery in many settings, very few centers will train the parents to store these sources. By information, we used the clinical material which is normally going to wastage. Preparation of Human umbilical cord samples was aseptically collected after delivery full-term by normal labor/C-section at Dept of obstetrics & gynaecology, Narayana Medical College Hospital and the protocol is approved by the Institutional Ethics Committee. After the delivery of the baby, the cord is collected in a sterile flask containing PBS supplemented with 100 U/mL penicillin, 0.1 mg/mL streptomycin, and 0.25 mg/mL amphotericin B. Cords were processed within 12 hr of birth in aseptic condition under Bio Safety Cabinet. Immediately, cords transferred to the laboratory and washed twice with PBS to remove remnant blood. The net weight of each cord was measured. Length of the cord is estimated. Cord is processed in a sterile 10 cm petridish; cord is divided into four sections according to its length from placenta to baby side. Each sectioned cord was spliced into 3–5 cm long pieces. Horizontal sections of each piece are engraved then cut open lengthwise with scissors and the outer sheath of umbilical cord sections peeled to extract its inner surface with WJ. The exposed vein and arteries removed by pulling with forceps and peeled away from the WJ. This tissue was chopped into small pieces placed into 50 ml centrifuge tube and digested with collagenase enzyme solution for 2 hrs at 37° C with constant agitation. The cord pieces are crushed using cell scrappers to disintegrate tissue into cell suspension. This cell suspension is moved to a new sterile 50ml centrifuge tube containing trypsin enzyme solution with constant agitation for 5minutes and stopped by adding Bovine serum albumin. After the second enzymatic digestion is complete, the cord pieces are scrapped with cell scrapper through 40micron filter to remove as many cells from Wharton’s jelly as possible. Cell suspension was centrifuged at 1000 rpm for 5 min and 10 ml of DMEM-F12 medium is added to the cell pellet. Cells were counted using a hemocytometer.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Collection of cord blood before placental delivery (in utero)</strong><br />\r\nThe cord was clamped immediately after the delivery and iodine swab done. The umbilical cord blood was collected from the vein by gravity in a sterile 100 ml collection tube containing layered with heparin. This process was performed by trained obstetric staffs. Cord blood was stored at 4°C until processing.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>After placental delivery (ex utero) collection</strong><br />\r\nImmediately after the placental delivery, the placenta is placed in a sterile tray with a central hole that allows the umbilical cord to hang down. Ubilical cord cleaned with an iodine swab, cord blood was collected from the umbilical vein in a sterile collection 100 ml tube layered with heparin and stored at 4°C until processing. For both methods, obstetric and clinical maternal chart contains route of delivery, gestation number, placental weight, sex and weight were recorded.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Mononuclear cell (MNC) separation</strong><br />\r\nA range of 55-80 ml of cord blood was collected in 100ml tubes with heparin. Cord blood subjected to density gradient centrifugation procedure using Histopaque 1077. Briefly, cord blood was diluted equal volume of PBS and layered on Histopaque in 50 ml. centrifuge tube and centrifuged at 850xg for 20 minutes. The mononuclear cell layer was separated, washed twice with PBS and resuspended in 1ml of PBS. The cells were counted using haemocytometer. The MNCs were subjected to Magnetic sorting.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Flow cytometry</strong><br />\r\nCord blood and WJ tissue samples were analyzed for the expression of CD34 and CD90 makers. The MNCs were stained with anti-human monoclonal antibodies i.e., FITC conjugated CD34 and PE-conjugated CD90 (BD Biosciences), after which they were kept in refrigerator at 4 °C for 20–30 minutes by flow cytometry analysis by BD FACS Canto II with reference to mouse isotypic Immunoglobulin as control. WJ samples were also analyzed as above procedure and the data was analyzed using BD FACS DIVA Software.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Immunomagnetic separation</strong><br />\r\nMNCs collected from cord blood subjected to Magnetic activated cell sorting (MACS) by using positive selection to CD34 phenotypic markers. According to the manufacturer’s instructions, 50 to 200 x 106 mononuclear cells in PBS and 0.5% BSA were incubated for 15 min at 4°C with a blocking reagent (human IgG) and simultaneously with anti-human CD34 antibody. After incubation, the cells were washed to remove unbound antibody and loaded in LS+ column separation. Labelled cells by positive selection were collected by flushing LS+ column with 5 ml DMEM medium. The cell viability was checked using Trypan blue dye exclusion assay and cell count performed by using a hemocytometer. CD90 positive cells were also collected using the same procedure.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Cell viability and enumeration</strong><br />\r\nThe stock (0.4 %) was diluted with PBS to 0.2 %. Cell suspension and 0.2 % trypan blue solutions were mixed at 1:1. Dilution factor is 5. The tip of the pipette is placedin the V-shaped groove on the hemacytometer to load the sample into the chamber (about 15 microliters) Capillary action will draw the fluid into the chamber. The sample is allowed to settle for 1- 2 minutes so that the cells stop drifting around the chamber and most will be in the same plane of focus. To avoid drying, the hemacytometer was placed in a petri dish containing a moistened filter paper. Total and viable cells were observed and counted in nine squares of hemacytometer. Live cell count divided by total cell count is Cell viability. Viable cell count / Quadrants counted x Dilution factor x Hemocytometer factor x Current volume (mL) = Viable cell yield. Here, hemocytometer factor is 10000.The correlaion between stem cell markers (CD34+ and CD90+) with neonatal parameters. Statistical test was performed by statistical software using SPSS 20.0 version. P value < 0.05 was considered significant.</p>"
},
{
"section_number": 3,
"section_title": "RESULTS",
"body": "<p><strong>Clinical characteristics</strong><br />\r\nData about umbilical cord (UC), and its cord blood were obtained from 12 vaginal and 8 caesarean sections. Eight cases were excluded from CB units before processing in both vaginal and caesarean sections. The cord blood collected ranged from 55-80 ml from 12 umbilical veins. Placental weight observed to be ranged from 418-540 grams. A total of 12 cords were collected and the mean weight per length of cords were of 1.2 g/cm (range 0.7–1.75 grams/cm). The clinical characteristics of Umbilical cord samples were mentioned in <a href=\"#Table-1\">Table 1</a>.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>CD34+ cells in Cord Blood and WJ tissue</strong><br />\r\nAfter density separation, Total MNCs obtained ranged from umbilical cord blood were 3.5 x 106 to 4.1 x 108 perml. The mononuclear cell purity observed as 68% from umbilical cord blood sample. After magnetic separation, the purity of cord blood CD34+ cells shows ~85%. The concentration of CD34+ cells sorted cells from umbilical cord blood mononuclear cells ranged from 2.42 x106 to 3.65×106. Cells show a mean of 94% viability after Histopaque density gradient separation of umbilical cord MNCs and WJ of umbilical cord blood.</p>\r\n\r\n<div id=\"Table-1\">\r\n<p><strong><a href=\"https://jabet.bsmiab.org/table/178-1521008794-table1/\">Table-1</a> Table 1. </strong>Characteristics of UC samples that generated MSCs</p>\r\n</div>\r\n\r\n<p> </p>\r\n\r\n<p><strong>CD90+ population from WJ</strong><br />\r\nCD90 cells were isolated from 12 numbers of cords and counted on a hemocytometer. Cell yield was calculated at 27×104 viable cells per centimeter length of cord (range 80- 469×103 cm–1 cord) the total mean cord length is 48 cm. Viability of CD90+ was observed as 92±3% (mean± SD (<a href=\"#Table-2\">Table 2</a>). The purity observed CD90+ cells were 96.4±1.4% after MACs sorting.</p>\r\n\r\n<div id=\"Table-2\">\r\n<p><strong><a href=\"https://jabet.bsmiab.org/table/178-1521008794-table2/\">Table-2</a> Table 2.</strong> CD90 phenotype marker sorting levels in various sources.</p>\r\n</div>\r\n\r\n<p> </p>\r\n\r\n<p><strong>CD34 and CD45 markers in cord blood and WJ</strong><br />\r\nThe mean percentage CD34+cell count was 1.02 ± 0.72 in cord blood in recovery. Hematopoetic stem cell [CD34+/CD45–] cell number observed as 35 – 250 cells in cord blood. Whereas there was negligible or nil expression of CD34+CD45 cell number observed in WJ samples i.e. 0.3 ± 0.3.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>CD34 and CD90 in cord blood and WJ</strong><br />\r\nThe mean percentage CD90+ cell count was 13.5 ± 2.72 in WJ tissue in recovery. These cells showed 70-80% positivity with CD90 not with CD34 and CD45. There was CD90+CD45–CD34– cell number 30-110 in observed cord blood. The mean percentage of CD90+ was 0.23 ±0.17 in cord blood [<a href=\"#figure1\">Figure 1</a>]. Correlation of CD34 and CD90 with neonatal variables in c-section delivery, there was an increased expression of CD34 and CD90 when compared to the vaginal delivery without significant difference in both cell counts. Baby’s gender doesn’t shown association with number of CD34 and CD90. The mean CD34+ and CD90+ cell counts between the two delivery groups of birth weight was compared using post hoc analysis, which showed that increased cell counts with increasing birth weight. Birth weight with CD34+ cell count showing correlation coefficient of 0.909 with p value 0.0001. Birth weight with CD90+ cell count showing correlation coefficient of 0.890 with p value 0.0001. Hence birth weight may be act as significant predictors of both CD34+ and CD90+ cell yield of the cord blood and WJ tissue.</p>\r\n\r\n<div id=\"figure1\">\r\n<figure class=\"image\"><img alt=\"\" height=\"868\" src=\"/media/article_images/2024/14/08/Figure_1.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 1. </strong>Flow cytometry analysis demonstrated that the cells were positive for CD90 and negligible/negative for CD34 and CD45 in wharton’s jelly and also positive for CD34 and negative for CD45 in cord blood.</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Correlation of CD34 and CD90 with neonatal variables</strong><br />\r\nIn c-section delivery, there was an increased expression of CD34 and CD90 when compared to the vaginal delivery without significant difference in both cell counts. Baby’s gender doesn’t shown association with number of CD34 and CD90. The mean CD34+ and CD90+ cell counts between the two delivery groups of birth weight was compared using post hoc analysis, which showed that increased cell counts with increasing birth weight. Birth weight with CD34+ cell count showing correlation coefficient of 0.909 with p value 0.0001. Birth weight with CD90+ cell count showing correlation coefficient of 0.890 with p value 0.0001. Hence birth weight may be act as significant predictors of both CD34+ and CD90+ cell yield of the cord blood and WJ tissue. </p>"
},
{
"section_number": 4,
"section_title": "DISCUSSION",
"body": "<p>The human umbilical cord contains distinct anatomical regions, two umbilical arteries, umbilical vein, cord lining and Wharton’s jelly. Each of these regions comprises fibroblast like MSCs. Cells isolated from arteries, vein, cord lining and wharton’s jelly have the capacity of plastic adherence, multipotent and differentiating ability to cell types such as osteoblasts, adipocytes, chondrocytes, hepatocytes, and neural and cardiac cells. They express markers like bone marrow derived MSC, negative for haematopoietic and macrophage markers. In the current study, WJ tissue derived cells shown positive to CD90 and negative to CD34 and CD45 markers respectively. It indicates WJ derived cells fulfill the mesenchymal stem cell lineage. Whereas, cord blood sample shown an opposite pattern of expression. CB shown positive for CD34 and negative or less expression of CD45. It indicates that CB derived cells are of hematipoetic lineage.<br />\r\nPrevious studies have succeeded in the establishment of correlation of the hematopoietic stem cell content with the neonatal, obstetric and other factors. The present study designed to characterize the CD34+ cell content (hematopoetic) and CD90+ (mesenchymal) cell content in cord blood and wharton’s jelly tissue of the same pregnant with respect to the major neonatal parameters. In this study, there was significant impact of birth weight on CD34+/CD45- hematopoetic stem cell content of the cord blood, which also been reported by other study was found to be supportive to the present observation [<a href=\"#r-17\">17</a>]. Though birth weight was correlated significantly with CD34+/CD90+ cell counts, the increasing gestational age doesn’t showing significant increase in CD34+/CD90+ cell counts in the current study. Babies with higher weight shown higher CD34 expression in cord blood and also higher CD90 in WJ derived cell counts. Mode of delivery doesn’t shown any influence on hematopoetic and mesenchymal stem cell yield in our study, which also been supported by the previous study [<a href=\"#r-18\">18</a>]. Whereas, another study observed more progenitor cell content especially CD34+ cell concentration in case of cesarean mode of delivery than the vaginal delivery mode [<a href=\"#r-19\">19</a>]. In the current study, it was an attempt to made or formulate the guidelines related to maternal and neonatal parameters to store the cord blood sample or tissues before collecting the cord blood or tissue in India. Limitation of this study was small sample size, hence the observations should be validated in long term prospective studies in the country.<br />\r\nUnlike embryonic stem cells, adult stem cell usage in research and therapeutics doesn’t have any controversy because it does not require the creation/destruction of an embryo for the production of adult stem cells. In recent years, there seems to be an unbounded interest concerning mesenchymal stem cells (MSCs). Reason due to this is there are some exciting characteristics of MSC, which includes long-term ex vivo proliferation, multi-lineage potential and immunomodulatory properties. Bone marrow has some limitations as MSCs sources, including mesenchymal stem cell low frequency, isolation procedure is painful and the decline in MSC characteristics with donor’s age [<a href=\"#r-20\">20</a>]. Kobayashi et al [<a href=\"#r-2\">2</a>] demonstrated that umbilical cord which is discarded during the birth may have an inexhaustible stem cells source and showed similar characteristics with the MSCs from bone marrow. MSCs from Wharton’s jelly tissue may serve as large, rapidly available and ethically noncontroversial and doesn’t induce teratomas [<a href=\"#r-3\">3</a>]. WJ tissue derived MSCs are more primitive MSC than other tissue sources and doesn’t express MHC class II (HLA-DR) antigens [<a href=\"#r-15\">15</a>]. During embryonic development, wharton’s jelly is derived from extra-embryonic mesoderm and partly from the embryonic mesoderm.<br />\r\nThe isolation of MSCs from individual cord regions including cord vein and the perivascular region of Wharton’s jelly and cord blood has been reported by many groups. However, very few/none of the studies or indeed any other studies have compared all cord regions of the same pregnant individual. We described the characterisation of MSCs obtained from each of the distinct regions carefully dissected from the same umbilical cord, providing a direct and relevant comparison of cells with the potential for tissue engineering and repair.</p>"
},
{
"section_number": 5,
"section_title": "CONCLUSION",
"body": "<p>In this study MSCs from four distinct regions of the same cord (artery, vein, Wharton’s jelly, and cord lining), in addition to cord blood cells from same individual from the whole cord, have been isolated and compared. The MSCs, which express CD90 from the richest site of the umbilical cord (Wharton’s jelly) can only be isolated using enzymatic digestion. Cells from whole cord and cells isolated from individual cord regions have potential as a useful source for obtaining promising cell populations for further study. The present study also suggests positive association between hematopoietic stem cell content of the cord blood and the neonatal factor especially birth weight. Mesenchymal stem cell content also has shown positive correlation with birth weight.</p>"
},
{
"section_number": 6,
"section_title": "ACKNOWLEDGEMENT",
"body": "<p>This research does not receive any fund from public or private organization. Authors acknowledge to Narayana Medical College & Hospitals for providing reagents and infrastructure.</p>"
},
{
"section_number": 7,
"section_title": "CONFLICT OF INTEREST",
"body": "<p>The author declares that no conflict of interest exists.</p>"
}
],
"figures": [
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/14/08/Figure_1.jpg",
"caption": "Figure 1. Flow cytometry analysis demonstrated that the cells were\r\npositive for CD90 and negligible/negative for CD34 and CD45 in\r\nwharton’s jelly and also positive for CD34 and negative for CD45 in\r\ncord blood.",
"featured": false
}
],
"authors": [
{
"id": 37,
"affiliation": [
{
"affiliation": "Head of Advanced Research Center, Asst. Professor, Department of Genetics & Stem cell Research, Narayana Medical College, Nellore, \r\nAndhrapradesh, India."
}
],
"first_name": "Mahaboob Vali",
"family_name": "Shaik",
"email": "drmahaboobvs@gmail.com",
"author_order": 1,
"ORCID": "https://orcid.org/0000-0003-4059-1709",
"corresponding": true,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "Dr. Mahaboob Vali Shaik, Head of Advanced Research Center, Asst. Professor, Laboratory of Genetics & Stem cell \r\nResearch, Narayana Medical College, Nellore, Andhrapradesh, India, Phone-9985335296, Email: drmahaboobvs@gmail.com",
"article": 21
},
{
"id": 38,
"affiliation": [
{
"affiliation": "Professor, Department of Obstetrics & Gynaecology, Narayana Medical College& Hospital, Nellore, Andhrapradesh, India."
}
],
"first_name": "Hymavathi",
"family_name": "K",
"email": null,
"author_order": 2,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 21
},
{
"id": 39,
"affiliation": [
{
"affiliation": "Director & Cardiologist, Narayana Medical Institutions, Nellore, Andhrapradesh, India."
}
],
"first_name": "Subrahmanyam",
"family_name": "G",
"email": null,
"author_order": 3,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 21
}
],
"views": 792,
"downloads": 126,
"references": [
{
"id": 217,
"serial_number": 1,
"pmc": null,
"reference": "D’Ippolito G, Schiller PC, Ricordi C, Roos BA, Howard GA. Age‐related osteogenic potential of mesenchymal stromal stem cells from human vertebral bone marrow. Journal of Bone and Mineral Research. 1999;14:1115-22.",
"DOI": null,
"article": 21
},
{
"id": 218,
"serial_number": 2,
"pmc": null,
"reference": "Kobayashi K, Kubota T, Aso T. Study on myofibroblast differentiation in the stromal cells of Wharton’s jelly: expression and localization of alpha-smooth muscle actin. Early Hum Dev. 1998;51:223-33.",
"DOI": null,
"article": 21
},
{
"id": 219,
"serial_number": 3,
"pmc": null,
"reference": "Fong CY, Chak LL, Biswas A, Tan JH, Gauthaman K, Chan WK, et al. Human Wharton’s jelly stem cells have unique transcriptome profiles compared to human embryonic stem cells and other mesenchymal stem cells. Stem Cell Reviews and Reports. 2011;7:1-16.",
"DOI": null,
"article": 21
},
{
"id": 220,
"serial_number": 4,
"pmc": null,
"reference": "Sarugaser R, Lickorish D, Baksh D, Hosseini MM, Davies JE. Human umbilical cord perivascular (HUCPV) cells: a source of mesenchymal progenitors. Stem cells. 2005;23:220-9.",
"DOI": null,
"article": 21
},
{
"id": 221,
"serial_number": 5,
"pmc": null,
"reference": "Wang L, Detamore MS. Insulin‐like growth factor‐I improves chondrogenesis of predifferentiated human umbilical cord mesenchymal stromal cells. Journal of Orthopaedic Research. 2009;27:1109-15.",
"DOI": null,
"article": 21
},
{
"id": 222,
"serial_number": 6,
"pmc": null,
"reference": "Pereira WC, Khushnooma I, Madkaikar M, Ghosh K. Reproducible methodology for the isolation of mesenchymal stem cells from human umbilical cord and its potential for cardiomyocyte generation. Journal of tissue engineering and regenerative medicine. 2008;2:394-9.",
"DOI": null,
"article": 21
},
{
"id": 223,
"serial_number": 7,
"pmc": null,
"reference": "Conconi MT, Burra P, Di Liddo R, Calore C, Turetta M, Bellini S, et al. CD105 (+) cells from Wharton’s jelly show in vitro and in vivo myogenic differentiative potential. International journal of molecular medicine. 2006;18:1089-96.",
"DOI": null,
"article": 21
},
{
"id": 224,
"serial_number": 8,
"pmc": null,
"reference": "Campard D, Lysy PA, Najimi M, Sokal EM. Native umbilical cord matrix stem cells express hepatic markers and differentiate into hepatocyte-like cells. Gastroenterology. 2008;134:833-48.",
"DOI": null,
"article": 21
},
{
"id": 225,
"serial_number": 9,
"pmc": null,
"reference": "Chen M-Y, Lie P-C, Li Z-L, Wei X. Endothelial differentiation of Wharton’s jelly–derived mesenchymal stem cells in comparison with bone marrow–derived mesenchymal stem cells. Experimental hematology. 2009;37:629-40.",
"DOI": null,
"article": 21
},
{
"id": 226,
"serial_number": 10,
"pmc": null,
"reference": "Karahuseyinoglu S, Cinar O, Kilic E, Kara F, Akay GG, Demiralp DÖ, et al. Biology of stem cells in human umbilical cord stroma: in situ and in vitro surveys. Stem cells. 2007;25:319-31.",
"DOI": null,
"article": 21
},
{
"id": 227,
"serial_number": 11,
"pmc": null,
"reference": "Fu YS, Cheng YC, Lin MYA, Cheng H, Chu PM, Chou SC, et al. Conversion of human umbilical cord mesenchymal stem cells in Wharton’s jelly to dopaminergic neurons in vitro: potential therapeutic application for Parkinsonism. Stem cells. 2006;24:115-24.",
"DOI": null,
"article": 21
},
{
"id": 228,
"serial_number": 12,
"pmc": null,
"reference": "Maleki M, Parivar K, Nabiyouni M, Yaghmaei P, Naji M. Induction of alpha-crystallins expression in umbilical cord mesenchymal stem cells. Iranian Journal of Ophthalmology. 2010;22:67-71.",
"DOI": null,
"article": 21
},
{
"id": 229,
"serial_number": 13,
"pmc": null,
"reference": "Wang HS, Hung SC, Peng ST, Huang CC, Wei HM, Guo YJ, et al. Mesenchymal stem cells in the Wharton’s jelly of the human umbilical cord. Stem cells. 2004;22:1330-7.",
"DOI": null,
"article": 21
},
{
"id": 230,
"serial_number": 14,
"pmc": null,
"reference": "La Rocca G, Anzalone R, Farina F. The Expression of CD68 in Human Umbilical Cord Mesenchymal Stem Cells: New Evidences of Presence in Non‐Myeloid Cell Types. Scandinavian journal of immunology. 2009;70:161-2.",
"DOI": null,
"article": 21
},
{
"id": 231,
"serial_number": 15,
"pmc": null,
"reference": "Troyer DL, Weiss ML. Concise Review: Wharton’s Jelly‐Derived Cells Are a Primitive Stromal Cell Population. Stem cells. 2008;26:591-9.",
"DOI": null,
"article": 21
},
{
"id": 232,
"serial_number": 16,
"pmc": null,
"reference": "Ma L, Feng X, Cui B, Law F, Jiang X, Yang L, et al. Human umbilical cord Wharton’s Jelly-derived mesenchymal stem cells differentiation into nerve-like cells. CHINESE MEDICAL JOURNAL-BEIJING-ENGLISH EDITION-. 2005;118:1987.",
"DOI": null,
"article": 21
},
{
"id": 233,
"serial_number": 17,
"pmc": null,
"reference": "Wen S-H, Zhao W-L, Lin P-Y, Yang K-L. Associations among birth weight, placental weight, gestational period and product quality indicators of umbilical cord blood units. Transfusion and Apheresis Science. 2012;46:39-45.",
"DOI": null,
"article": 21
},
{
"id": 234,
"serial_number": 18,
"pmc": null,
"reference": "Lim FT, Van Winsen L, Willemze R, Kanhai HH, Falkenburg JH. Influence of delivery on numbers of leukocytes, leukocyte subpopulations, and hematopoietic progenitor cells in human umbilical cord blood. Blood Cells. 1994;20:547-58.",
"DOI": null,
"article": 21
},
{
"id": 235,
"serial_number": 19,
"pmc": null,
"reference": "Aroviita P, Teramo K, Hiilesmaa V, Kekomäki R. Cord blood hematopoietic progenitor cell concentration and infant sex. Transfusion. 2005;45:613-21.",
"DOI": null,
"article": 21
},
{
"id": 236,
"serial_number": 20,
"pmc": null,
"reference": "Chen MY, Lie PC, Li ZL, Wei X. Endothelial differentiation of Wharton’s jelly-derived mesenchymal stem cells in comparison with bone marrow-derived mesenchymal stem cells. Experimental hematology. 2009;37:629-40.",
"DOI": null,
"article": 21
}
]
},
{
"id": 15,
"slug": "178-1519046049-lipopolysaccharide-tolerance-attenuates-inflammatory-responses-by-increasing-heme-oxygenase-1-and-tristetraprolin-expression-in-raw2647-macrophages",
"featured": false,
"slider": false,
"issue": "Vol1 Issue1",
"type": "original_article",
"manuscript_id": "178-1519046049",
"recieved": "2018-01-13",
"revised": null,
"accepted": "2018-01-30",
"published": "2018-01-31",
"pdf_file": "https://jabet.bsmiab.org/media/pdf_file/2023/51/178-1519046049.pdf",
"title": "Lipopolysaccharide tolerance attenuates inflammatory responses by increasing heme oxygenase-1 and tristetraprolin expression in Raw264.7 macrophages",
"abstract": "<p>Inflammation is associated with the progression of a variety of diseases. Lipopolysaccharides (LPS) tolerance is recognized to reduce proinflammatory responses. Heme oxygenase1 (HO1) and tristetraprolin (TTP) are induced by LPS tolerance and mediate the anti-inflammatory effects. However, it was not clear whether two molecules are linked in LPS tolerance. In this study, we sought to determine whether HO1 associates with TTP to mediate the anti-inflammatory effects of LPS tolerance. LPS treatment significantly increased mRNA and protein level of HO1 and TTP in a time dependant fashion while LPS significantly decreased mRNA and protein level of TNFα in Raw264.7 macrophages. LPS tolerance inhibited TNFα mRNA and protein while HO1 and TTP level was still increased. In HO1 deficient macrophages, LPS tolerance failed to attenuate TNFα mRNA expression but TTP level was still decreased. Our results suggest that HO1 and TTP are functionally linked in mediating anti-inflammatory effects of LPS tolerance. This novel LPS tolerance-HO1-TTP signaling pathway provides new possibilities for the treatment of inflammatory diseases.</p>",
"journal_reference": "J Adv Biotechnol Exp Ther. 2018; 1(1) : 07-10.",
"academic_editor": "Md Nabiul Islam, PhD;\r\nYamaguchi University, Japan.",
"cite_info": "Moni A, Uddin MJ. Lipopolysaccharide tolerance attenuates inflammatory responses by increasing heme oxygenase-1 and tristetraprolin expression in Raw264.7 macrophages. J Adv Biotechnol Exp Ther. 2018; 1(1) : 07-10.",
"keywords": [
"LPS tolerance",
"anti-inflammatory effects",
"HO1",
"TTP"
],
"DOI": "10.5455/jabet.d5",
"sections": [
{
"section_number": 1,
"section_title": "INTRODUCTION",
"body": "<p style=\"text-align:justify\">Inflammation is associated with the progression of a variety of diseases such as cancer, atherosclerosis, asthma, psoriasis, and so on. Lipopolysaccharides (LPS), secreted by gram-negative bacteria activates macrophages. Activated macrophages mediate excessive production of proinflammatory cytokines including tumor necrosis factor α (TNFα), which leads to serious systemic disorders. Pretreatment of LPS is involved with reduction of sensitivity to subsequent challenge of LPS which is termed LPS tolerance. LPS tolerance was observed in vivo as well as in vitro with a reduced production of inflammatory cytokines in response to a secondary LPS stimulation. However, the precise mechanisms involved in LPS tolerance-induced alteration of the proinflammatory responses remains unclear. </p>\r\n\r\n<p>The inflammatory response modulated by post-transcriptional control [<a href=\"#r-1\">1</a>, <a href=\"#r-2\">2</a>] is dependent on AU-rich element (ARE)–mediated mechanisms [<a href=\"#r-3\">3</a>-<a href=\"#r-5\">5</a>]. The ARE-binding protein, tristetraprolin (TTP) binds and destabilizes pro-inflammatory mRNAs including TNFα [<a href=\"#r-6\">6</a>-<a href=\"#r-9\">9]</a>. The importance of TTP in inhibiting inflammatory responses has been implicated in TTP-deficient mice that develop severe inflammatory arthritis, autoimmune dysfunction, and myeloid hyperplasia [<a href=\"#r-10\">10</a>]. These mice produce increasing amount of the proinflammatory cytokine TNFα by macrophages and suffer from slow growth, underweight and the development of polyarticular arthritis. In mouse macrophages, increase in TTP expression and activity was associated with decrease expression of TNFα mRNA [<a href=\"#r-11\">11</a>]. In addition, increase in TTP mRNA expression in response to LPS stimulation was observed in dendritic cells.</p>\r\n\r\n<p>It was reported that LPS treatment induces the expression of TTP [<a href=\"#r-12\">12</a>, <a href=\"#r-13\">13</a>]. LPS-induced TTP seems to exert an inhibitory function against LPS-induced lung inflammation, because TTP deficiency aggravates LPS-induced lung inflammation.</p>\r\n\r\n<p>Hemeoxygenase 1 (HO1) is a stress-inducible protein and the anti-inflammatory effects of HO1 may have therapeutic potential in inflammatory conditions such as arthritis [<a href=\"#r-14\">14</a>]and inflammatory bowel disease [<a href=\"#r-15\">15</a>]. HO1 and TTP are functionally linked in mediating the anti-inflammatory effects of nicotine; HO1 is necessary for the induction of TTP. Increased HO1 expression was observed in the lung during LPS tolerance and cross tolerance [<a href=\"#r-16\">16</a>]. In addition, overexpression of hepatic HO1 has been observed during LPS tolerance [<a href=\"#r-17\">17</a>]. Currently, there are no reports regarding the effects of HO1 on the regulation of TTP expression on LPS tolerance under pro-inflammatory conditions.</p>\r\n\r\n<p>Therefore, in our study, we focused on detailed mechanisms under laying the LPS tolerance effects on HO1- TTP expression in macrophages.</p>"
},
{
"section_number": 2,
"section_title": "MATERIALS AND METHODS",
"body": "<p><strong>Reagents and antibodies</strong></p>\r\n\r\n<p>Anti-β-actin, anti-TTP, anti-HO1 and horseradish peroxidase conjugated secondary (anti-mouse IgG and anti-goat IgG) antibodies and small interfering RNA (siRNAs) against mouse HO1, and control siRNA were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). LPS and protease inhibitor cocktail sets were purchased from Sigma Aldrich (St. Louis, MO, USA). Dulbecco’s Modified Egale Medium (DMEM), fetal bovine serum (FBS), penicillin–streptomycin and sodium pyruvate were purchased from Invitrogen (Grand Island, NY, USA). All other chemicals were purchased from Sigma-Aldrich.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Cell culture</strong></p>\r\n\r\n<p>Mouse Raw264.7 macrophage cells were cultured in DMEM supplemented with 10% FBS and 1% penicillin-streptomycin. Cells were seeded in 6-well plates at a density of 5×105/ml in DMEM medium supplemented with 10% FBS. Supernatants were collected.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Transfection</strong></p>\r\n\r\n<p>Cells (5×105/ml) were cultured in 6-well plate for 18h and transfected with HO1 siRNA (100 nM) using Lipofectamine 2000 (Invitrogen, CA, USA). Cells were pretreated with LPS 100 ng/ml for 24 h in the presence or absence of 1 μg/ml LPS for 6 h. The expression levels of HO1, TTP, or TNFα mRNA were analyzed by q-RT-PCR.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Western blot assays</strong></p>\r\n\r\n<p>Cell lysates were prepared using RIPA buffer containing protease inhibitors and phosphatase inhibitors and total protein concentration of the lysates was measured using a BCA Protein Assay kit (Pierce Biotechnology Inc., Rockford, IL, USA). Proteins were resolved by SDS-PAGE, transferred onto polyvinylidenedifluoride (PVDF) membrane, and probed with appropriate dilutions of the following antibodies: anti-TTP, anti-HO1, and anti-β-actin. Immunoreactivity was detected using the ECL detection system (GE healthcare Bio-Sciences Corp, NJ). Films were exposed at multiple time points to ensure that the images were not saturated.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Reverse transcription-polymerase chain reaction (RT-PCR)</strong></p>\r\n\r\n<p>Total RNA was extracted using TRIzol reagent (Invitrogen) according to manufacturer’s instructions. DNase I-treated total RNA (2 μg) was reversed transcribed using M-MLV reverse transcriptase (Promega Corporation, WI, USA) and oligo-dT (Promega Corporation, WI, USA). Semi-quantitative RT-PCR was performed using Taq polymerase (Solgent, Daejeon, Korea) and PCR primer pairs as follows: GAPDH: 5′-aggccggtgctgagtatgtc-3′, 5′-tgcctgcttcaccttct-3′; HO-1: 5′-tcccagacaccgctcctccag-3′, 5′-ggatttggggctggtttc-3′; TTP: 5′-ctctgccatctacgagagcc-3′, 5′-gatggagtccgagtttatgttcc-3′; TNFα, 5′-agcccacgtcgtagcaaaccaccaa-3′, 5′-acacccattcccttcacagagcaat-3′. The gene amplification reaction conditions were as follows: denaturation at 94ºC for 0.5 min; annealing at 58-62ºC (based on the melting temperature of each respective primer) for 0.5 min; extension at 72ºC for 1 min: PCR cycles were determined according to a kinetic profile. GAPDH was used as an internal loading control.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Enzyme linked immunosorbent assay (ELISA)</strong></p>\r\n\r\n<p>TNFa in the cell supernatants were analyzed using Mouse TNF-alpha Quantikine ELISA Kit (R&D Systems).</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Statistical analysis</strong></p>\r\n\r\n<p>Statistical differences between groups were evaluated by one-way ANOVA or student’s t-test. A p value of <0.05 was considered statistically significant.</p>"
},
{
"section_number": 3,
"section_title": "RESULTS",
"body": "<p><strong>LPS increases HO1 and TTP expression in macrophages</strong><br />\r\nHO1 is a well known anti-inflammatory protein and TTP is known to destabilization of TNFα mRNA ARE sites. To prepare LPS-induced endotoxin tolerance in vitro model we treated Raw264.7 macrophages with LPS in a time dependent manner. LPS treatment significantly increased mRNA and protein level of HO1 (<a href=\"#figure1\">Figure 1A, C</a> and <a href=\"#figure1\">F</a>) and TTP (<a href=\"#figure1\">Figure 1A, D</a> and<a href=\"#figure1\"> F</a>) in a time dependant fashion. Conversely, LPS significantly decreased mRNA and protein level of TNFα in a time dependant manner (<a href=\"#figure1\">Figure 1A, B</a> and<a href=\"#figure1\"> E</a>). These events were more significant at the time of 18 or 24 h.</p>\r\n\r\n<div id=\"figure1\">\r\n<figure class=\"image\"><img alt=\"\" height=\"294\" src=\"/media/article_images/2024/59/08/Figure_1.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 1. </strong>LPS increases HO1 and TTP expression in Raw264.7 macrophages. (A-F) Cells were treated with LPS (100 ng/ml) in time dependant manner (0, 3, 6, 9, 18, and 24 h. (A-D) The levels of TNFα, HO-1, and TTP were measured by semi-qRT-PCR. ImageJ software was used for densitometry analysis. Values are mean ± SEM (n = 3). The representative bands are shown (A). (E) Cell supernatants were analyzed for TNFα by ELISA. Values are mean ± SD (n = 3). *p<0.05. (F) The protein levels of HO1 and TTP were analyzed by western blotting. The representative bands are shown.</figcaption>\r\n</figure>\r\n</div>\r\n\r\n<p> </p>\r\n\r\n<p><strong>LPS tolerance inhibits inflammatory responses via HO1-TTP expression in macrophages</strong><br />\r\nTo determine whether LPS tolerance has any effect on TTP expression as well as inflammatory responses. We pretreated Raw264.7 macrophages with LPS for 24 h (1<sup>st</sup> hit as LPS tolerant model) and again stimulated with LPS for 6 h (2<sup>nd</sup> hit). Results showed that LPS tolerance prevented effect of 2<sup>nd</sup> challenge of LPS as measured by TNFα mRNA and protein (<a href=\"#figure2\">Figure 2 A</a> and <a href=\"#figure2\">C</a>) while HO1 and TTP level was still increased (<a href=\"#figure2\">Figure 2 A</a> and <a href=\"#figure2\">B</a>). To further confirm the effect of TTP in LPS tolerance, we used HO1 siRNA. In con siRNA, LPS tolerance limited effects of 2<sup>nd</sup> hit of LPS as shown by TNFα mRNA expression (<a href=\"#figure2\">Figure 2D</a>) but in HO1 siRNA, LPS tolerance failed to suppress TNFα mRNA expression (<a href=\"#figure2\">Figure 2D</a>) while HO1 siRNA effectively suppressed HO1 and TTP expression which indicating the link between HO1 and TTP as described previously. Evidences and our data indicate that LPS tolerance in macrophages is mediated by HO1-TTP axis.</p>\r\n\r\n<div id=\"figure2\">\r\n<figure class=\"image\"><img alt=\"\" height=\"381\" src=\"/media/article_images/2024/59/08/Figure_2.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 2.</strong> LPS tolerance inhibits inflammatory responses via TTP expression in Raw264.7 macrophages. (A) Cells were pretreated with LPS (100 ng/ml) for 24 h and then, challenged with LPS (1 ug/ml) for 6 h. (A). The levels of TTP, HO1, and TNFα mRNA were analyzed by RT-PCR. (B) The levels of TTP, and HO1 proteins were analyzed by western blotting. (C) Cell supernatants were analyzed for TNFα protein by ELISA. Values are mean ± SD (n = 3), *p<0.05. (D) Cells were transfected with HO1 siRNA, and then cells were pretreated with LPS (100 ng/ml) for 24 h and challenged with LPS (1ug/ml) for another 6 h. The levels of TTP, HO-1, and TNFα mRNA were analyzed by RT-PCR.</figcaption>\r\n</figure>\r\n</div>"
},
{
"section_number": 4,
"section_title": "DISCUSSION",
"body": "<p>Endotoxin tolerance induced by LPS preconditioning is recognized to reduce proinflammatory responses. In our study, we reported that LPS tolerance attenuates inflammatory responses through increasing HO1-TTP system.</p>\r\n\r\n<p>Molecular mechanisms for LPS tolerance have long been investigated [<a href=\"#r-18\">18</a>]. LPS-signaling pathways have been shown to be affected by several proteins. In addition to HO1, TTP is induced by LPS treatment [<a href=\"#r-13\">13</a>]. HO1 and TTP provide protection against inflammatory diseases and LPS tolerance was found to increase their expression [<a href=\"#r-13\">13</a>, <a href=\"#r-17\">17</a>]. Thus, it is hypothesized that LPS tolerance increases the expression of HO1 and TTP and thus, suppresses inflammatory responses. TTP suppresses inflammation by destabilizing the mRNAs of pro-inflammatory cytokines. LPS-induced TTP seems to exert an inhibitory function against LPS-induced lung inflammation, because TTP deficiency aggravates LPS-induced lung inflammation. HO1 mediates its anti-inflammatory effects through TTP [<a href=\"#r-19\">19</a>]. Despite evidence for the roles of HO1 and TTP in LPS tolerance-mediated anti-inflammatory effects, the functional links between HO1 and TTP in endtoxin tolerance state is unclear. In this report, we demonstrate that increased expression of HO1 and TTP functions in the endtoxin tolerance -mediated anti-inflammatory signaling pathway. Our data provide evidence for the requirement of HO1 in LPS tolerance-induced upregulation of TTP in macrophages. In addition, silencing of HO1 blocks the effect of LPS tolerance on the induction of TTP and inhibits LPS-induced TNFα production both in vitro. Collectively, our data suggest that HO1 and TTP mediate the anti-inflammatory function of LPS tolerance in the HO1-TTP signaling pathway.</p>\r\n\r\n<p>In summary, these results suggest that HO1 and TTP are functionally linked in mediating the anti-inflammatory function of LPS tolerance. LPS tolerance enhances the expression of HO1-TTP levels which in turn inhibits LPS-induced TNFa production. This study identified a novel LPS tolerance-HO1-TTP signaling pathway responsible for the inhibition of LPS-driven inflammation and potentially provides the rationale for novel therapeutic approaches for the management of inflammatory diseases.</p>"
},
{
"section_number": 5,
"section_title": "ACKNOWLEDGEMENT",
"body": "<p>The author thanks to Dr. Md Jamal Uddin for his kind help during experiments and manuscript preparation.</p>"
},
{
"section_number": 6,
"section_title": "CONFLICT OF INTERESTS",
"body": "<p style=\"text-align:justify\">The author declares no conflict of interest.</p>"
}
],
"figures": [
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/59/08/Figure_1.jpg",
"caption": "Figure 1. LPS increases HO1 and TTP expression in Raw264.7 macrophages. (A-F) Cells were treated with LPS (100 ng/ml) in time dependant manner (0, 3, 6, 9, 18, and 24 h. (A-D) The levels of TNFα, HO-1, and TTP were measured by semi-qRT-PCR. ImageJ software was used for densitometry analysis. Values are mean ± SEM (n = 3). The representative bands are shown (A). (E) Cell supernatants were analyzed for TNFα by ELISA. Values are mean ± SD (n = 3). *p<0.05. (F) The protein levels of HO1 and TTP were analyzed by western blotting. The representative bands are shown.",
"featured": true
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/59/08/Figure_2.jpg",
"caption": "Figure 2. LPS tolerance inhibits inflammatory responses via TTP expression in Raw264.7 macrophages. (A) Cells were pretreated with LPS (100 ng/ml) for 24 h and then, challenged with LPS (1 ug/ml) for 6 h. (A). The levels of TTP, HO1, and TNFα mRNA were analyzed by RT-PCR. (B) The levels of TTP, and HO1 proteins were analyzed by western blotting. (C) Cell supernatants were analyzed for TNFα protein by ELISA. Values are mean ± SD (n = 3), *p<0.05. (D) Cells were transfected with HO1 siRNA, and then cells were pretreated with LPS (100 ng/ml) for 24 h and challenged with LPS (1ug/ml) for another 6 h. The levels of TTP, HO-1, and TNFα mRNA were analyzed by RT-PCR.",
"featured": false
}
],
"authors": [
{
"id": 12,
"affiliation": [
{
"affiliation": "ABEx Bio-Research Center, Azampur, Dakkhinkhan, Dhaka-1230, Bangladesh"
}
],
"first_name": "Akhi",
"family_name": "Moni",
"email": "akhimoni840818@gmail.com",
"author_order": 1,
"ORCID": "https://orcid.org/0000-0002-5812-7755",
"corresponding": true,
"co_first_author": true,
"co_author": false,
"corresponding_author_info": "Akhi Moni, PhD; ABEx Bio-Research, 3161, East Azampur, Imam Bukhari (RA) Road, Azampur, Dakkhinkhan, Dhaka 1230, Bangladesh. e-mail: akhimoni840818@gmail.com",
"article": 15
},
{
"id": 13,
"affiliation": [
{
"affiliation": "ABEx Bio-Research Center, Azampur, Dakkhinkhan, Dhaka-1230, Bangladesh"
},
{
"affiliation": "Graduate School of Pharmaceutical Sciences, College of Pharmacy, Ewha Womans University, Seoul, Korea"
}
],
"first_name": "Md Jamal",
"family_name": "Uddin",
"email": "hasan800920@gmail.com",
"author_order": 2,
"ORCID": "https://orcid.org/0000-0003-2911-3255",
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 15
}
],
"views": 545,
"downloads": 133,
"references": [
{
"id": 20,
"serial_number": 1,
"pmc": null,
"reference": "Stoecklin G, Anderson P. Posttranscriptional mechanisms regulating the inflammatory response. AdvImmunol. 2006;89:1-37.",
"DOI": null,
"article": 15
},
{
"id": 21,
"serial_number": 2,
"pmc": null,
"reference": "Hollams EM, Giles KM, Thomson AM, Leedman PJ. MRNA stability and the control of gene expression: implications for human disease. Neurochem Res. 2002;27:957-80.",
"DOI": null,
"article": 15
},
{
"id": 68,
"serial_number": 3,
"pmc": null,
"reference": "Clark A. Post-transcriptional regulation of pro-inflammatory gene expression. Arthritis Res. 2000;2:172-4.",
"DOI": null,
"article": 15
},
{
"id": 69,
"serial_number": 4,
"pmc": null,
"reference": "Kracht M, Saklatvala J. Transcriptional and post-transcriptional control of gene expression in inflammation. Cytokine. 2002;20:91-106.",
"DOI": null,
"article": 15
},
{
"id": 70,
"serial_number": 5,
"pmc": null,
"reference": "Khabar KS. The AU-rich transcriptome: more than interferons and cytokines, and its role in disease. J Interferon Cytokine Res. 2005;25:1-10.",
"DOI": null,
"article": 15
},
{
"id": 71,
"serial_number": 6,
"pmc": null,
"reference": "Carballo E, Lai WS, Blackshear PJ. Feedback inhibition of macrophage tumor necrosis factor-alpha production by tristetraprolin. Science. 1998;281:1001-5.",
"DOI": null,
"article": 15
},
{
"id": 72,
"serial_number": 7,
"pmc": null,
"reference": "Phillips K, Kedersha N, Shen L, Blackshear PJ, Anderson P. Arthritis suppressor genes TIA-1 and TTP dampen the expression of tumor necrosis factor alpha, cyclooxygenase 2, and inflammatory arthritis. Proc Natl Acad Sci U S A. 2004;101:2011-6.",
"DOI": null,
"article": 15
},
{
"id": 73,
"serial_number": 8,
"pmc": null,
"reference": "Park D, Lee EK, Jang EJ, Jeong HO, Kim BC, Ha YM, et al. Identification of the dichotomous role of age-related LCK in calorie restriction revealed by integrative analysis of cDNA microarray and interactome. Age (Dordr).",
"DOI": null,
"article": 15
},
{
"id": 74,
"serial_number": 9,
"pmc": null,
"reference": "Qian X, Ning H, Zhang J, Hoft DF, Stumpo DJ, Blackshear PJ, et al. Posttranscriptional regulation of IL-23 expression by IFN-gamma through tristetraprolin. J Immunol.186:6454-64.",
"DOI": null,
"article": 15
},
{
"id": 75,
"serial_number": 10,
"pmc": null,
"reference": "Taylor GA, Carballo E, Lee DM, Lai WS, Thompson MJ, Patel DD, et al. A pathogenetic role for TNF alpha in the syndrome of cachexia, arthritis, and autoimmunity resulting from tristetraprolin (TTP) deficiency. Immunity. 1996;4:445-54.",
"DOI": null,
"article": 15
},
{
"id": 76,
"serial_number": 11,
"pmc": null,
"reference": "Anderson P. Post-transcriptional control of cytokine production. Nat Immunol. 2008;9:353-9.",
"DOI": null,
"article": 15
},
{
"id": 77,
"serial_number": 12,
"pmc": null,
"reference": "Brooks SA, Connolly JE, Rigby WF. The role of mRNA turnover in the regulation of tristetraprolin expression: evidence for an extracellular signal-regulated kinase-specific, AU-rich element-dependent, autoregulatory pathway. J Immunol. 2004;172:7263-71.",
"DOI": null,
"article": 15
},
{
"id": 78,
"serial_number": 13,
"pmc": null,
"reference": "Chen YL, Jiang YW, Su YL, Lee SC, Chang MS, Chang CJ. Transcriptional regulation of tristetraprolin by NF-kappaB signaling in LPS-stimulated macrophages. Mol Biol Rep. 2013;40:2867-77.",
"DOI": null,
"article": 15
},
{
"id": 79,
"serial_number": 14,
"pmc": null,
"reference": "Devesa I, Ferrandiz ML, Terencio MC, Joosten LA, van den Berg WB, Alcaraz MJ. Influence of heme oxygenase 1 modulation on the progression of murine collagen-induced arthritis. Arthritis Rheum. 2005;52:3230-8.",
"DOI": null,
"article": 15
},
{
"id": 80,
"serial_number": 15,
"pmc": null,
"reference": "Hegazi RA, Rao KN, Mayle A, Sepulveda AR, Otterbein LE, Plevy SE. Carbon monoxide ameliorates chronic murine colitis through a heme oxygenase 1-dependent pathway. J Exp Med. 2005;202:1703-13.",
"DOI": null,
"article": 15
},
{
"id": 81,
"serial_number": 16,
"pmc": null,
"reference": "Koch A, Boehm O, Zacharowski PA, Loer SA, Weimann J, Rensing H, et al. Inducible nitric oxide synthase and heme oxygenase-1 in the lung during lipopolysaccharide tolerance and cross tolerance. Crit Care Med. 2007;35:2775-84.",
"DOI": null,
"article": 15
},
{
"id": 82,
"serial_number": 17,
"pmc": null,
"reference": "Soriano RN, Ravanelli MI, Batalhao ME, Carnio EC, Branco LG. Glucocorticoids downregulate systemic nitric oxide synthesis and counteract overexpression of hepatic heme oxygenase-1 during endotoxin tolerance. Can J Physiol Pharmacol. 2013;91:861-5.",
"DOI": null,
"article": 15
},
{
"id": 83,
"serial_number": 18,
"pmc": null,
"reference": "Ziegler-Heitbrock HW. Molecular mechanism in tolerance to lipopolysaccharide. J Inflamm. 1995;45:13-26.",
"DOI": null,
"article": 15
},
{
"id": 84,
"serial_number": 19,
"pmc": null,
"reference": "Jamal Uddin M, Joe Y, Zheng M, Blackshear PJ, Ryter SW, Park JW, et al. A functional link between heme oxygenase-1 and tristetraprolin in the anti-inflammatory effects of nicotine. Free Radic Biol Med. 2013;65:1331-9.",
"DOI": null,
"article": 15
}
]
},
{
"id": 18,
"slug": "178-1514390013-role-of-drug-metabolic-enzymes-and-transporters-in-drug-drug-interactions-between-antiretroviral-and-antituberculosis-drugs",
"featured": false,
"slider": false,
"issue": "Vol1 Issue1",
"type": "review_article",
"manuscript_id": "178-1514390013",
"recieved": "2017-12-25",
"revised": null,
"accepted": "2018-01-16",
"published": "2018-01-25",
"pdf_file": "https://jabet.bsmiab.org/media/pdf_file/2023/53/178-1514390013.pdf",
"title": "Role of drug metabolic enzymes and transporters in drug-drug interactions between antiretroviral and antituberculosis drugs",
"abstract": "<p>Drug-drug interactions is one of the major determinant in drug development and clinical applications. Individual differences in pharmacokinetics may cause extensive variability in drug efficacy, toxicity and adverse drug reactions, and represent a major concern in drug development. According to world health organization, an estimated 1.2 million patients were diagnosed with HIV/TB coinfection in 2015, and approximately 400,000 coinfected individuals were died in 2016. During treatment of tuberculosis and human immunodeficiency virus (HIV) coinfections, a combination of multiple therapeutic drugs is used, which is potential for interactions between coadministered drugs. Drug metabolizing enzymes and transporters plays vital role in the disposition, pharmacokinetics as well as efficacy and toxicity of many antituberculosis (anti-TB) and antiretroviral (ARVs) drugs. The current understanding of drug-drug interactions involving the first-line antituberculosis and antiretroviral (ARVs) drugs are reviewed and summarized in this article emphasizing the role of drug transporters and metabolic enzymes. It is found that several clinical drug-interactions are caused by the phase I and II metabolic enzymes (cytochrome-P-450s, CYPs, UDP-glucuronosyltransferases, UGTs) and uptake/ efflux transporters, particularly permeability glycoprotein, brest cancer resistance protein, multidrug resistance protein and solute carrier transporters). Overall, this study will be helpful to understand the drug-interactions between the ARV or anti-TB drug regimen or with concomitantly used drugs at a glance. By understanding the unique functions and characteristics of enzymes and transporters, it may use in the management of drug interactions in the treatment of HIV and TB.</p>",
"journal_reference": "J Adv Biotechnol Exp Ther. 2018; 1(1) : 17-24.",
"academic_editor": "Dr. Md Ataur Rahman, Korea Institute of Science and Technology, South Korea.",
"cite_info": "Parvez MM, Kaisar N, Shin JG. Role of drug metabolic enzymes and transporters in drug-drug interactions between antiretroviral and antituberculosis drugs. J Adv Biotechnol Exp Ther. 2018; 1(1) : 17-24.",
"keywords": [
"Membrane Transporters",
"Antiretroviral and antituberculosis drugs.",
"Drug-drug interactions",
"Metabolic Enzymes"
],
"DOI": "10.5455/jabet.d2",
"sections": [
{
"section_number": 1,
"section_title": "INTRODUCTION",
"body": "<p>To date, it is very common to alteration of drug’s clinical response in multiple drug regimen by another drug is defined as a drug-drug interactions (DDI). The alterations of response may result from the change in drug disposition which is believed to cause by induction or inhibition of transporters or metabolic enzymes and drug transporters play role in drug absorption, distribution, metabolism and excretion (ADME), those DDIs are known for pharmacokinetic interactions [<a href=\"#r-1\">1</a>]. Drug transporters belonging to two major super-families of membrane-associated proteins, the solute carriers (SLCs), which primarily function as a cellular drug influx mechanism, and the ATP-binding cassette (ABC) transporters which in contrast, efflux drugs/ compounds from cellular targets, are known to play a vital role in overall drug disposition (i.e., drug distribution to target tissues/cells and drug clearance from the liver and kidney). These transporters are also thought to be contributor in pharmacokinetic drug-drug interactions, clinical response as well as toxicity [<a href=\"#r-2\">2</a>][<a href=\"#r-3\">3</a>]. On the other hand, drug metabolizing enzymes comprising of the Phase I oxidative enzymes (cytochrome-P-450s, CYPs) and the conjugation Phase II enzymes (sulfotransferases, glucuronyl transferases, <em>N</em>-acetyltransferases and glutathione-S-transferases), are key players in exogenous/ endogenous compound and drug metabolism, and are differentially expressed in various mammalian tissues. The pharmacokinetic DDIs are responsible for approximately 20-30% of the adverse drug reactions in general population, they also account for about 10% of the cases under emergency department and contribute 3-5% of the medication errors in-patients [<a href=\"#r-4\">4</a>][<a href=\"#r-5\">5</a>]. Since the drug pharmacokinetics can be significantly altered by both transporters and enzyme mediated DDIs, it can potentially affect to the therapeutic efficacy or toxicity of a drug [<a href=\"#r-6\">6</a>]. Generally, DDIs based on metabolism are due to induction and/or inhibition of metabolic enzymes (CYP450s or UGTs), are considered as most prominent cause [<a href=\"#r-7\">7</a>]. Based on the therapeutic and toxic effects of both the parent drug and its metabolites, the clinical consequences depends on the CYP450 enzyme induction or inhibition and this may be particularly significant to those drugs has narrow therapeutic window because DDIs based on metabolism may cause changes in the concentration of drug up to 10 fold whose biotransformation is induced or inhibited [<a href=\"#r-8\">8</a>]. To date, drug-metabolizing enzymes mediated DDIs have been extensively studied <em>in vitro</em> as well as <em>in vivo</em>, emerging understanding has led to uptake and efflux transporters being recognized as a significant determinant of drug disposition that may cause significant DDIs. Passive diffusion through the membrane has been viewed as dominant in the disposition of most drugs, but it is now well recognized that carrier-mediated transport plays a vital role [<a href=\"#r-9\">9</a>]<br />\r\nAcquired Immunodeficiency syndrome (AIDS) caused by Human Immunodeficiency Virus 1 (HIV-1) is considered as a major public health issue globally. In 2016, an estimated 1.2 million people died of AIDS-related illness out of 36.7 million people affected with this disease worldwide [<a href=\"#r-10\">10</a>]. Although, the combination of antiretroviral therapy (cART) has effectively decreased HIV-1 related mortality and morbidity, some opportunistic infections are increasingly common and causing death of HIV patients. For example, tuberculosis (TB) is one of the leading opportunistic infections and cause of death in HIV patients. According to World Health Organization, an estimated 1.2 million patients were diagnosed with HIV/TB coinfection in 2015, and approximately 400,000 coinfected individuals were died in 2016 (World Health Organization, 2015, 2016b). In Europe, HIV/TB coinfection increased by 40% over the last 5 years [<a href=\"#r-12\">12</a>]. Overall, the risk of developing TB is reported to be 26–31 times higher in HIV-infected compared to HIV non-infected persons [<a href=\"#r-13\">13</a>]. Similarly, the most common adverse events such as psychiatric disorders (i.e., depression and anxiety) are found in HIV infected patients than non-infected. The prevalence of depression and anxiety were reported to be 22-50% and 2-40%, respectively in HIV infected individuals globally [<a href=\"#r-14\">14</a>-<a href=\"#r-15\">15</a>], whereas serious mental illness (SMI) in HIV infected individuals was estimated to be between 3% to 23%, a 10 fold higher occurrence than in the healthy population in United States (0.4%) [<a href=\"#r-16\">16</a>-<a href=\"#r-18\">18</a>].<br />\r\nThe concurrent use of antiretroviral (ARV) and antituberculosis (anti-TB) drugs have significantly reduced the mortality rate of HIV/TB patients [<a href=\"#r-19\">19</a>]. However, the use of multiple drug regimens in a combination are known to produce drug–drug interactions and overlapping toxicities which are the most challenging aspects of managing both diseases. According to recent guidelines, at least one ARV regimen from the integrase strand transfer inhibitors (INSTIs) class (Douletegravir/Elvitegravir/Raltegravir) and two from the nucleotide reverse transcriptase inhibitors (NRTIs) class (abacavir/lamivudine/tenofovir/emtricitabine) are recommended as a first line therapy for HIV infection (<a href=\"#Table-1\">Table 1</a>) [<a href=\"#r-20\">20</a>]. In addition protease inhibitors (PIs) (Ritonavir/Atazanavir) are recommended as a boosting therapy depending on treatment outcomes [<a href=\"#r-20\">20</a>]. Whereas, a combined regimen of rifampicin, isoniazid, ethambutol and pyrazinamide are recommended as a first line therapy for tuberculosis (TB) in the world [<a href=\"#r-10\">10</a>]. HIV-infected individuals are often suffered with opportunistic infections, and are likely to be prescribed with multiple medications to manage complex medical conditions in addition to their ART regimens, making them vulnerable to the effects of potential and serious drug-drug interactions, caused by alteration in drug transport and metabolism affecting pharmacokinetic/pharmacodynamic (PK/PD) properties.</p>\r\n\r\n<div id=\"Table-1\">\r\n<p><strong><a href=\"https://jabet.bsmiab.org/table/178-1514390013-table1/\">Table-1</a>Table 1: </strong>Recommended antiretroviral regimen for initial therapy.<strong> </strong></p>\r\n</div>"
},
{
"section_number": 2,
"section_title": "DISCUSSION",
"body": "<p><strong>Drug Metabolizing Enzymes (DMEs)</strong><br />\r\nDrug-metabolizing reactions are mainly classified into phase I and phase II metabolism. Briefly, phase I reactions involve oxidation, reduction or hydrolysis of the drug, and are primarily mediated by the cytochrome P450 (CYP450) family of enzymes. Phase II reactions involve covalently binding of an endogenous compound, most often glucuronide acid, glutathione or sulphate, to the phase I metabolite. For example, phase II enzymes are uridine diphosphoglucuronosyl transferases (UGTs) and glutathione-S-transferases (GSTs). The phase II conjugation produces a more polar metabolite and promotes excretion of the drug from the tissue, normally via efflux transporters. The CYP450 enzymes responsible for metabolism of xenobiotics are primarily expressed in the liver and intestines, and to less extend in the lung, kidney and central nervous system. CYP450 enzymes are generally located in the endoplasmic reticulum of cells where they carry out the nicotinamide adenine dinucleotide phosphate-oxidase (NADPH)-dependent oxidation of a diversity of substrates. The superfamily of CYP450 enzymes comprises 57 genes which have been organized into families (denoted by the first identification number, e.g. CYP3) and subfamilies (denoted by letters, e.g. CYP3A) [<a href=\"#r-21\">21</a>]. The individual isoenzymes within each subfamily are further denoted by numbers, e.g. CYP3A4. CYP450 enzymes are in general promiscuous in their capacity to bind and metabolite substrates, and thus, there is significant overlapping substrate specificity among CYP enzymes. At present, among the human CYP450 isoenzymes, CYP1A1, CYP1A2, CYP2B6, CYP2C9, CYP2C19, CYP2D6, CYP2E1, CYP3A4 and CYP3A5 are considered of particular importance in drug metabolism (<a href=\"#Table-2\">Table 2</a>) [<a href=\"#r-22\">22</a>]. To date, it is hypothesized that the biotransformation of approximately 40-50% of all xenobiotic drugs on the market are caused by CYP3A-mediated oxidation [<a href=\"#r-23\">23</a>]. For the majority of individuals, the human CYP3A isoform CYP3A4 are the most abundant CYP enzyme expressed in the liver and intestine, and considered as the major drug-metabolizing enzyme in human [<a href=\"#r-24\">24</a>]<sup>,</sup>[<a href=\"#r-25\">25</a>].</p>\r\n\r\n<div id=\"Table-2\">\r\n<p><strong><a href=\"https://jabet.bsmiab.org/table/178-1514390013-table2/\">Table-2</a> Table 2. </strong>Major Drug Metabolizing Human CytochromeP450 Enzymes in Human. </p>\r\n</div>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Drug Transporters (DTs)</strong><br />\r\nAt present drug transporters are mainly classified into two major gene superfamily, the solute carrier (SLC) superfamily and the ATP-binding cassette (ABC) superfamily [9]. The members of the SLC superfamily are facilitated transporters or ion-coupled secondary transporters, frequently associated with uptake of compounds from the blood into tissues or organs such as the liver and kidney, or in the absorption from the gastrointestinal tract into the systemic circulation [<a href=\"#r-26\">26</a>]. The SLC superfamily include 48 subfamilies of which the most investigated are the organic anion transporting polypeptides (OATPs, SLCO), the organic cation transporters (OCTs) and organic anion transporters (OATs), as well as the more recently identified multidrug and toxin extrusion (MATE, SLC47A) family which is involved in drug excretion from the kidney and liver (<a href=\"#Table-3\">Table 3</a>) [<a href=\"#r-27\">27</a>].</p>\r\n\r\n<div id=\"Table-3\">\r\n<p><a href=\"https://jabet.bsmiab.org/table/178-1514390013-table3/\"><strong>Table-3.</strong></a> <strong>Table 3. </strong>Major ADMET Drug Transporters in Human. </p>\r\n</div>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Interactions of ARVs and/ or Anti-TB Drugs with Membrane Transporters and Metabolic Enzymes</strong><br />\r\nSeveral first line ARV therapy drugs, such as integrase strand transfer inhibitors (INSTIs) (raltegravir, dolutegravir and elvitegravir), nucleoside reverse transcriptase inhibitors (NRTIs) (lamivudine, abacavir, tenofovir, emtricitabine) and protease inhibitors (PIs) (atazanavir, ritonavir) are substrates of ABC efflux transporters such as P-glycoprotein (P-gp); multidrug resistance proteins (MRPs) and/or breast cancer resistance protein (BCRP), and thus could potentially interfere with the ABC transporter function of effluxing drugs from their cellular targets. In addition, metabolic enzymes such as members of the CYP450 family (e.g., CYP3A4) and UDP-Glucuronosyl transferase family (e.g., UGT1A1) can also contribute to an extensive intracellular degradation of several ARVs, particularly protease inhibitors and integrase strand transfer inhibitors [<a href=\"#r-28\">28</a>]. Together, these effects can significantly contribute to poor tissue and intracellular ARVs concentrations [<a href=\"#r-29\">29</a>]. Recently, this has been clearly demonstrated the atazanavir disposition is significantly involved with transporters in a P-gp/BCRP knockout mouse [<a href=\"#r-30\">30</a>]. In particular, significantly higher atazanavir brain and testicular tissue concentrations was found among the P-gp knockout compared to the wild type mice suggesting that the efflux transporters localized at the blood-brain barrier and blood-testicular barrier are important determinants of this HIV protease inhibitor tissue distribution [<a href=\"#r-30\">30</a>]. In addition, several anti-TB drugs are known to be substrates and potent modulators of drug transporters (influx and efflux) and metabolic enzymes (both phase I and II enzymes), and are implicated in many clinical drug- interactions [<a href=\"#r-31\">31</a>-<a href=\"#r-33\">33</a>]. For example, among the first line anti-TB drug regimen, rifampicin is known as a substrate, inducer and inhibitor of liver enzymes (CYP450s & UGTs), and P-gp, OATPs (OATP1B1, -1B3) and implicated in the reduction of many concomitant drugs (statins<strong>) </strong>[<a href=\"#r-31\">31</a>]<sup>,</sup>[<a href=\"#r-34\">34</a>]<sup>,</sup>[<a href=\"#r-35\">35</a>]<sup>,</sup>[<a href=\"#r-36\">36</a>]. Similarly, ethambutol is another first line anti-TB drug undergoes non-cyp450 mediated metabolism and substrate/inhibitor of multiple OCT transporters [<a href=\"#r-37\">37</a>]<sup>,</sup>[<a href=\"#r-38\">38</a>]<sup>,</sup>[<a href=\"#r-39\">39</a>].</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Complexities of Drug-Drug Interactions Involving Anti-HIV and Anti-TB/Psychotic Drugs</strong><br />\r\nThe combination of HIV and TB often is referred to as a syndemic, where two or more diseases interact in a synergistic fashion, and leads to an overall increased burden of both diseases [<a href=\"#r-40\">40</a>]. One associated factor for such occurrence could the use of multiple drug regimens for the treatment of HIV co-infection with TB, and the overlapping drug-drug interactions (DDI) involving drug transport and metabolism, which may result in toxic and/or sub-therapeutic drug levels leading to therapy change or treatment failure. Several DDIs involving anti-HIV and anti-TB drugs have been reported previously. For example, in healthy volunteers raltegravir AUC was increased by 77% when coadministered with rifapentine and this DDIs was poorly understood [<a href=\"#r-41\">41</a>]. On the other hand, rifampin (rifampicin analogue) decreased raltegravir concentration by up to 60% among healthy subjects [<a href=\"#r-42\">42</a>]. It is known that, raltegravir primary metabolize by UGT1A1 and actively transported by P-gp and BCRP could play role in this DDIs. Another integrase inhibitor, Dolutegravir is primarily metabolized by UGT1A1 and CYP3A4 (lesser extent) [<a href=\"#r-43\">43</a>] and also a potent inhibitor of OCT2 as well as substrate of P-gp and BCRP [<a href=\"#r-44\">44</a>]. In a phase-1 clinical study, rifampin was reported to decrease dolutegravir AUC approximately 54% whereas rifabutin showed no significant effect on dolutegravir pharmacokinetics [<a href=\"#r-45\">45</a>]. However, dolutegravir is a potent inhibitor of organic cation transporter 2 (OCT2) and multidrug and toxic compound extrusion (MATE) transporters, and metformin is transported by OCTs transporter for disposition. As a result, metformin higher plasma concentration was found when coadministered with dolutegravir among healthy subjects. Regarding the first line therapy ARV drugs, particularly NRTI that has been studied with raltegravir is the tenofovir. In a study among healthy subjects, the plasma concentrations of raltegravir was increased by 49% due to a yet unexplained mechanism which needs further investigation to understand the exact mechanism of this clinical interactions [<a href=\"#r-46\">46</a>]. In another study, raltegravir concentrations was reported to increase by 72% when combinedly administered with atazanavir in healthy subjects [<a href=\"#r-47\">47</a>]. Similarly, dolutegravir AUC was increased approximately 91% and 62% (when boosted with ritonavir) by atazanavir in healthy subjects [<a href=\"#r-43\">43</a>]. Whereas, atazanavir is known to inhibit UGT1A1 enzyme, P-gp and BCRP efflux transporters [<a href=\"#r-48\">48</a>] responsible for metabolism and transport of raltegravir and dolutegravir [<a href=\"#r-49\">49</a>][<a href=\"#r-44\">44</a>]. It was proposed that, inhibition of enzymes and transporters by atazanavir could play role in the substantial increase of raltegravir/dolutegravir plasma concentration. In addition, atazanavir is known to increase tenofovir AUC by 21% in healthy subjects [<a href=\"#r-50\">50</a>]. The MRP2, a renal efflux pump could play role in this drug interactions which potentially could lead to renal toxicity [<a href=\"#r-51\">51</a>]. This potential MRP2 mediated drug interactions remains unclear and further investigations would be helpful to understand. These complexity in drug interactions can be overcome by extensive mechanistic data for metabolizing enzymes and transporters.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Overlapping Toxicities with ARVs and Anti-TB Drug</strong> <strong>Regimen</strong><br />\r\nOverlapping toxicities and adverse reactions are common when multiple ARVs and anti-TB drugs are co-administered in a given time [<a href=\"#r-52\">52</a>] (<a href=\"#Table-4\">Table 4</a>) (<a href=\"#figure1\">Figure 1</a>). For example, Isoniazid, stauvudine and didanosine are well-known for drug induced neuropathy [<a href=\"#r-53\">53</a>]. As a result, increased hepatoxicity when nonnucleoside reverse transcriptase inhibitors, (NNRTIs, particularly nevirapine) were prescribed with anti-TB regimens was remains as a concerns for adverse event [<a href=\"#r-54\">54</a>]. In another study, found an increased hepatotoxicity in efavirenz [<a href=\"#r-54\">54</a>]<sup>,</sup>[<a href=\"#r-55\">55</a>] and nevirapine [<a href=\"#r-54\">54</a>] co-administration with anti-TB regimens in 30% HIV/TB patients [<a href=\"#r-56\">56</a>]. Again, in case of lopinavir [<a href=\"#r-57\">57</a>], atazanavir [<a href=\"#r-58\">58</a>] and saquinavir [<a href=\"#r-59\">59</a>] boosting with ritonavir in combination with rifampin caused severe hepatotoxicity in healthy subjects. It seemed, most common hepatotoxicity was reported among the subjects when protease inhibitors (PI) boosted with rifampin. In regard to sequence of drug therapy may play a role and hepatoxic metabolite of PIs comes from the rifampin induced CYP450s or ritonavir may cause reduction in metabolic clearance was hypothesized[<a href=\"#r-58\">58</a>].</p>\r\n\r\n<div id=\"Table-4\">\r\n<p><a href=\"https://jabet.bsmiab.org/table/178-1514390013-table4/\"><strong>Table 4. </strong></a><strong>Table 4. </strong>Overlapping ping side effect profiles of first-line antituberculosis drugs and antiretroviral drugs.</p>\r\n</div>\r\n\r\n<div id=\"figure1\">\r\n<figure class=\"image\"><img alt=\"\" height=\"361\" src=\"/media/article_images/2024/29/08/Figure1.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 1.</strong> Involvement of drug metabolizing enzymes and transporters in pharmacokinetics. This figure represents the involvement of metabolic enzymes and transporters can produce drug-drug interaction by inhibition/ induction of the corresponding enzymes and transporters in the different tissues (GI, Liver, Brain and Kidney). </figcaption>\r\n</figure>\r\n</div>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Transporter-Enzyme Interplay in Drug Disposition</strong><br />\r\nAt this advance science of age, a considerable overlap in substrate specificity and tissue distribution among membrane transporters and metabolizing enzymes were found throughout the body [60]. Recently, a significant substrate overlap between hepatic uptake transporters and enzymes has been recognized, [<a href=\"#r-60\">60</a>] e.g. between CYP3A4 and OATPs [<a href=\"#r-61\">61</a>]. However, the interactive nature of CYP3A and multidrug resistance protein 1 (MDR1) is the most extensively studied interplay between enzymes and transporters [<a href=\"#r-60\">60</a>]<sup>,</sup>[<a href=\"#r-62\">62</a>]<sup>,</sup>[<a href=\"#r-63\">63</a>]. CYP3A and MDR1 act as a coordinated barrier for xenobiotics. [<a href=\"#r-64\">64</a>] However, several clinical studies explained that the role of intestinal MDR1 greatly played role by limiting absorption of the drug [<a href=\"#r-62\">62</a>]<sup>,</sup>[<a href=\"#r-63\">63</a>]<sup>,</sup>[<a href=\"#r-64\">64</a>]. In the intestine, where the drug enters the enterocytes from the luminal side, MDR1 majorly play role in the process of the drug reaching to the enzyme through repeated cycles of absorption and efflux, giving CYP3A mediated metabolism prior to enter into systemic circulation. This phenomenon may explained as the drug from the enterocytes return to the lumen without metabolism by P-gp mediated efflux or other apical efflux transporters thereby allowing re-entry into enterocytes and increased chance of metabolic conversion by CYP3A [<a href=\"#r-65\">65</a>]. Again, in the liver, the drug enters into the hepatocytes from the basolateral side and encounters CYP3A prior to MDR1-mediated efflux into the bile canaliculi, drugs will not re-enter the cells because of concentration gradient, as result limited metabolism occurs and more parent drug cross the membrane.</p>"
},
{
"section_number": 3,
"section_title": "CONCLUSIONS",
"body": "<p>Drug-drug interactions is common phenomena in multidrug regimen, especially in TB/ HIV treatment. The metabolic enzymes and transport proteins are differentially expressed in several tissues and play a crucial role in the drugs disposition and clinical response. The emphasis on metabolic enzymes and drugs transporters are well given since a couple of years and has been attracted by many to investigate on them for their clinical drug-interactions. After having several clinical mechanistic studies, it is believed that, several drugs are influenced by in the interplay between metabolic enzymes and drug transporters. Also, still limited data are available to conclude or characterize precisely the DDIs, there is a requirement of more mechanistic understanding of several complex drug-interactions between antituberculosis and antiretroviral drugs. This indicates the requirement of more mechanistic studies to understand clinical pharmacokinetic/ pharmacodynamic DDIs. Furthermore, base-poke studies on drug metabolic enzymes and transporters role in the disposition of the drug/s will be helpful to optimize drug-dose regimen in pharmacotherapy to HIV-TB.</p>"
},
{
"section_number": 4,
"section_title": "ACKNOWLEDGEMENT",
"body": "<p>We are thankful to Md. Tozammel Hoque for excellent opinion on scientific aspects of this article. We declare no conflicts of interest or other relevant affiliations, financial involvement, or agreement/interest with any organization or governing body in this study. Additionally, there was no technical assistance in preparation of this manuscript.</p>"
},
{
"section_number": 5,
"section_title": "AUTHOR CONTRIBUTIONS",
"body": "<p>SS performed the experiment; MNI and MEH conceived the study; SS and MNI analyzed the data; SS and MNI. wrote the paper.</p>"
},
{
"section_number": 6,
"section_title": "CONFLICT OF INTERESTS",
"body": "<p>The authors declare no conflict of interest.</p>"
}
],
"figures": [
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/29/08/Figure1.jpg",
"caption": "Figure 1. Involvement of drug metabolizing enzymes and transporters in pharmacokinetics. This figure represents the involvement of metabolic enzymes and transporters can produce drug-drug interaction by inhibition/ induction of the corresponding enzymes and transporters in the different tissues (GI, Liver, Brain and Kidney).",
"featured": true
}
],
"authors": [
{
"id": 29,
"affiliation": [
{
"affiliation": "Department of Pharmacology and PharmacoGenomics Research Center, Inje University College of Medicine, Busan, Republic of Korea."
},
{
"affiliation": "Department of Pharmaceutical Sciences, Leslie Dan Faculty of Pharmacy, University of Toronto, Canada."
}
],
"first_name": "Md Masud",
"family_name": "Parvez",
"email": "masud.parvez@utoronto.ca",
"author_order": 1,
"ORCID": "https://scholar.google.com/citations?user=45gf75QAAAAJ&hl=en",
"corresponding": true,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "Md Masud Parvez, PhD; Department of Pharmacology and PharmacoGenomics Research Center, Inje University College of Medicine, Busan, Republic of Korea and Department of Pharmaceutical Sciences, Leslie Dan Faculty of Pharmacy, University of Toronto, \r\nCanada.. e-mail: masud.parvez@utoronto.ca",
"article": 18
},
{
"id": 30,
"affiliation": [
{
"affiliation": "Department of Pharmacology and PharmacoGenomics Research Center, Inje University College of Medicine, Busan, Republic of Korea."
}
],
"first_name": "Nazia",
"family_name": "Kaisar",
"email": null,
"author_order": 2,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": true,
"corresponding_author_info": "",
"article": 18
},
{
"id": 31,
"affiliation": [
{
"affiliation": "Department of Pharmacology and PharmacoGenomics Research Center, Inje University College of Medicine, Busan, Republic of Korea."
},
{
"affiliation": "Department of Clinical Pharmacology, Inje University Busan Paik Hospital, Busan, Republic of Korea"
}
],
"first_name": "Jae-Gook",
"family_name": "Shin",
"email": null,
"author_order": 3,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": true,
"corresponding_author_info": "",
"article": 18
}
],
"views": 1100,
"downloads": 95,
"references": [
{
"id": 125,
"serial_number": 1,
"pmc": null,
"reference": "Köhler GI, Bode-Böger SM, Busse R, Hoopmann M, Welte T, Böger RH. Drug-drug interactions in medical patients: effects of in-hospital treatment and relation to multiple drug use. Int J Clin Pharmacol Ther. 2000; 38: 504–13.",
"DOI": null,
"article": 18
},
{
"id": 126,
"serial_number": 2,
"pmc": null,
"reference": "König J, Müller F, Fromm MF. Transporters and Drug-Drug Interactions: Important Determinants of Drug Disposition and Effects. Pharmacol Rev. 2013; 65: 944-66.",
"DOI": null,
"article": 18
},
{
"id": 127,
"serial_number": 3,
"pmc": null,
"reference": "Zanger UM, Schwab M. Cytochrome P450 enzymes in drug metabolism: Regulation of gene expression, enzyme activities, and impact of genetic variation. Pharmacol Ther. 2013; 138: 103–41.",
"DOI": null,
"article": 18
},
{
"id": 128,
"serial_number": 4,
"pmc": null,
"reference": "Hohl CM, Dankoff J, Colacone A, Afilalo M. Polypharmacy, adverse drug-related events, and potential adverse drug interactions in elderly patients presenting to an emergency department. Ann Emerg Med. 2001; 38: 666–71.",
"DOI": null,
"article": 18
},
{
"id": 129,
"serial_number": 5,
"pmc": null,
"reference": "Leape LL, Bates DW, Cullen DJ, Cooper J, Demonaco HJ, Gallivan T et al. Systems analysis of adverse drug events. ADE Prevention Study Group. Jama. 1995; 274: 35–43.",
"DOI": null,
"article": 18
},
{
"id": 130,
"serial_number": 6,
"pmc": null,
"reference": "Leape LL, Bates DW, Cullen DJ, Cooper J, Demonaco HJ, Gallivan T et al. Systems analysis of adverse drug events. ADE Prevention Study Group. Jama. 1995; 274: 35–43.",
"DOI": null,
"article": 18
},
{
"id": 131,
"serial_number": 7,
"pmc": null,
"reference": "Pirmohamed M. Cytochrome P450 enzyme polymorphisms and adverse drug reactions. Toxicology. 2003; 192: 23–32.",
"DOI": null,
"article": 18
},
{
"id": 132,
"serial_number": 8,
"pmc": null,
"reference": "Teo YL, Ho HK, Chan A. Metabolism-related pharmacokinetic drug-drug interactions with tyrosine kinase inhibitors: Current understanding, challenges and recommendations. Br J Clin Pharmacol. 2015; 79: 241–53.",
"DOI": null,
"article": 18
},
{
"id": 133,
"serial_number": 9,
"pmc": null,
"reference": "Consortium IT, Giacomini K, Huang S-M, Tweedie D, Benet L, Brouwer K et al. Membrane transporters in drug development. Nat Rev Drug Discov. 2010; 9: 215–36.",
"DOI": null,
"article": 18
},
{
"id": 134,
"serial_number": 10,
"pmc": null,
"reference": "World Health Organization. Global Health Observatory (GHO) data: Global situation and trends of HIV. 2016; http://www.who.int/gho/hiv/en/.",
"DOI": null,
"article": 18
},
{
"id": 135,
"serial_number": 11,
"pmc": null,
"reference": "World Health Organization. Tuberculosis and HIV. 2016; http://www.who.int/hiv/topics/tb/about_tb/en/.",
"DOI": null,
"article": 18
},
{
"id": 136,
"serial_number": 12,
"pmc": null,
"reference": "Gao J, Zheng P, Fu H. Prevalence of TB/HIV Co-Infection in Countries Except China: A Systematic Review and Meta-Analysis. PLoS One. 2013; 8.",
"DOI": null,
"article": 18
},
{
"id": 137,
"serial_number": 13,
"pmc": null,
"reference": "World Health Organization (WHO). Tuberculosis and HIV. http://www.who.int/hiv/topics/tb/about_tb/en/.",
"DOI": null,
"article": 18
},
{
"id": 138,
"serial_number": 14,
"pmc": null,
"reference": "C O-LBSLSEA. HIV infection and psychiatric illness. African J Psychiatry (South Africa). 2009; 12: 115–28.",
"DOI": null,
"article": 18
},
{
"id": 139,
"serial_number": 15,
"pmc": null,
"reference": "Del Guerra FB, Fonseca JLI, Figueiredo VM, Ziff EB, Konkiewitz EC. Human immunodeficiency virus-associated depression: Contributions of immuno-inflammatory, monoaminergic, neurodegenerative, and neurotrophic pathways. J. Neurovirol. 2013; 19: 314–27.",
"DOI": null,
"article": 18
},
{
"id": 140,
"serial_number": 16,
"pmc": null,
"reference": "Lee S, Rothbard AB, Noll E, Blank MB. Use of HIV and psychotropic medications among persons with serious mental illness and HIV/AIDS. Adm Policy Ment Health. 2011; 38: 335–44.",
"DOI": null,
"article": 18
},
{
"id": 141,
"serial_number": 17,
"pmc": null,
"reference": "Cournos F, McKinnon K. HIV seroprevalence among people with severe mental illness in the United States: A critical review. Clin. Psychol. Rev. 1997; 17: 259–69.",
"DOI": null,
"article": 18
},
{
"id": 142,
"serial_number": 18,
"pmc": null,
"reference": "Rabkin JG. HIV and depression: 2008 Review and update. Curr. HIV/AIDS Rep. 2008; 5: 163–71.",
"DOI": null,
"article": 18
},
{
"id": 143,
"serial_number": 19,
"pmc": null,
"reference": "Abdool Karim SS, Naidoo K, Grobler A, Padayatchi N, Baxter C, Gray AL et al. Integration of antiretroviral therapy with tuberculosis treatment. N Engl J Med. 2011; 365: 1492–501.",
"DOI": null,
"article": 18
},
{
"id": 144,
"serial_number": 20,
"pmc": null,
"reference": "AIDS info. Guidelines for the Use of Antiretroviral Agents in Adults and Adolescents Living with HIV. 2017.",
"DOI": null,
"article": 18
},
{
"id": 145,
"serial_number": 21,
"pmc": null,
"reference": "Nelson DR, Zeldin DC, Hoffman SMG, Maltais LJ, Wain HM, Nebert DW. Comparison of cytochrome P450 (CYP) genes from the mouse and human genomes, including nomenclature recommendations for genes, pseudogenes and alternative-splice variants. Pharmacogenetics. 2004; 14: 1–18.",
"DOI": null,
"article": 18
},
{
"id": 146,
"serial_number": 22,
"pmc": null,
"reference": "Guengerich FP, Martin M V., Beaune PH, Kremers P, Wolff T, Waxman DJ. Characterization of rat and human liver microsomal cytochrome P-450 forms involved in nifedipine oxidation, a prototype for genetic polymorphism in oxidative drug metabolism. J Biol Chem. 1986; 261: 5051–60.",
"DOI": null,
"article": 18
},
{
"id": 147,
"serial_number": 23,
"pmc": null,
"reference": "Thummel KE, Wilkinson GR. In vitro and in vivo drug interactions involving human CYP3A. Annu Rev Pharmacol Toxicol. 1998; 38: 389–30.",
"DOI": null,
"article": 18
},
{
"id": 148,
"serial_number": 24,
"pmc": null,
"reference": "Guengerich FP. Cytochrome P-450 3A4: regulation and role in drug metabolism. Annu Rev Pharmacol Toxicol. 1999; 39: 1–17.",
"DOI": null,
"article": 18
},
{
"id": 149,
"serial_number": 25,
"pmc": null,
"reference": "Paine MF, Hart HL, Ludington SS, Haining RL, Rettie AE, Zeldin DC. The human intestinal cytochrome P450 ‘pie’. Drug Metab Dispos. 2006; 34: 880–86.",
"DOI": null,
"article": 18
},
{
"id": 150,
"serial_number": 26,
"pmc": null,
"reference": "Lin L, Yee SW, Kim RB, Giacomini KM. SLC transporters as therapeutic targets: Emerging opportunities. Nat. Rev. Drug Discov. 2015; 14: 543–60.",
"DOI": null,
"article": 18
},
{
"id": 151,
"serial_number": 27,
"pmc": null,
"reference": "Roth M, Obaidat A, Hagenbuch B. OATPs, OATs and OCTs: The organic anion and cation transporters of the SLCO and SLC22A gene superfamilies. Br. J. Pharmacol. 2012; 165: 1260–87.",
"DOI": null,
"article": 18
},
{
"id": 152,
"serial_number": 28,
"pmc": null,
"reference": "Kis O, Sankaran-Walters S, Hoque MT, Walmsley SL, Dandekar S, Bendayan R. HIV-1 Alters Intestinal Expression of Drug Transporters and Metabolic Enzymes: Implications for Antiretroviral Drug Disposition. Antimicrob Agents Chemother. 2016; 60: 2771–81.",
"DOI": null,
"article": 18
},
{
"id": 153,
"serial_number": 29,
"pmc": null,
"reference": "Alam C, Whyte-Allman SK, Omeragic A, Bendayan R. Role and modulation of drug transporters in HIV-1 therapy. Adv Drug Deliv Rev 2016; 103: 121–43.",
"DOI": null,
"article": 18
},
{
"id": 154,
"serial_number": 30,
"pmc": null,
"reference": "Robillard KR, Chan GNY, Zhang G, la Porte C, Cameron W, Bendayan R. Role of P-Glycoprotein in the Distribution of the HIV Protease Inhibitor Atazanavir in the Brain and Male Genital Tract. Antimicrob Agents Chemother. 2014; 58: 1713–22.",
"DOI": null,
"article": 18
},
{
"id": 155,
"serial_number": 31,
"pmc": null,
"reference": "Shimokawa Y, Yoda N, Kondo S, Yamamura Y, Takiguchi Y, Umehara K. Inhibitory Potential of Twenty Five Anti-tuberculosis Drugs on CYP Activities in Human Liver Microsomes. Biol Pharm Bull. 2015; 38: 1425–29.",
"DOI": null,
"article": 18
},
{
"id": 156,
"serial_number": 32,
"pmc": null,
"reference": "Cao L, Greenblatt DJ, Kwara A. Inhibitory Effects of Selected Antituberculosis Drugs on Common Human Hepatic Cytochrome P450 and UDP-glucuronosyltransferase Enzymes. Drug Metab Dispos. 2017; 45: 1035–43.",
"DOI": null,
"article": 18
},
{
"id": 157,
"serial_number": 33,
"pmc": null,
"reference": "Te Brake LHM, Russel FGM, Van Den Heuvel JJMW, De Knegt GJ, De Steenwinkel JE, Burger DM et al. Inhibitory potential of tuberculosis drugs on ATP-binding cassette drug transporters. Tuberculosis. 2016; 96: 150–57.",
"DOI": null,
"article": 18
},
{
"id": 158,
"serial_number": 34,
"pmc": null,
"reference": "Bolt HM. Rifampicin, A Keystone Inducer of Drug Metabolism: From Herbert Remmer’s Pioneering Ideas to Modern Concepts. Drug Metab Rev. 2004; 36: 497–09.",
"DOI": null,
"article": 18
},
{
"id": 159,
"serial_number": 35,
"pmc": null,
"reference": "Glaeser H, Drescher S, Eichelbaum M, Fromm MF. Influence of rifampicin on the expression and function of human intestinal cytochrome P450 enzymes. Br J Clin Pharmacol. 2005; 59.",
"DOI": null,
"article": 18
},
{
"id": 160,
"serial_number": 36,
"pmc": null,
"reference": "Ma X, Cheung C, Krausz KW, Shah YM, Wang T, Idle JR et al. A double transgenic mouse model expressing human pregnane X receptor and cytochrome P450 3A4. Drug Metab Dispos. 2008; 36: 2506–12.",
"DOI": null,
"article": 18
},
{
"id": 161,
"serial_number": 37,
"pmc": null,
"reference": "Te Brake LHM, Van Den Heuvel JJMW, Buaben AO, Van Crevel R, Bilos A, Russel FG et al. Moxifloxacin is a potent in vitro inhibitor of OCT- and MATE-mediated transport of metformin and ethambutol. Antimicrob Agents Chemother. 2016; 60: 7105–114.",
"DOI": null,
"article": 18
},
{
"id": 162,
"serial_number": 38,
"pmc": null,
"reference": "Pan X, Wang L, Gründemann D, Sweet DH. Interaction of ethambutol with human organic cation transporters of the SLC22 family indicates potential for drug-drug interactions during antituberculosis therapy. Antimicrob Agents Chemother. 2013; 57: 5053–59.",
"DOI": null,
"article": 18
},
{
"id": 163,
"serial_number": 39,
"pmc": null,
"reference": "Parvez MM, Kaisar N, Shin HJ, Jung JA, Shin JG. Inhibitory Interaction Potential of 22 Antituberculosis Drugs on Organic Anion and Cation Transporter of SLC22A family. Antimicrob Agents Chemother. 2016; 60: 6558–67.",
"DOI": null,
"article": 18
},
{
"id": 164,
"serial_number": 40,
"pmc": null,
"reference": "Kwan C, Ernst JD. HIV and tuberculosis: A deadly human syndemic. Clin. Microbiol. Rev. 2011; 24: 351–76.",
"DOI": null,
"article": 18
},
{
"id": 165,
"serial_number": 41,
"pmc": null,
"reference": "Weiner M, Egelund EF, Engle M, Kiser M, Prihoda TJ, Gelfond JAL et al. Pharmacokinetic interaction of rifapentine and raltegravir in healthy volunteers. J Antimicrob Chemother. 2014; 69: 1079–85.",
"DOI": null,
"article": 18
},
{
"id": 166,
"serial_number": 42,
"pmc": null,
"reference": "Wenning LA, Hanley WD, Brainard DM, Petry AS, Ghosh K, Jin B et al. Effect of rifampin, a potent inducer of drug-metabolizing enzymes, on the pharmacokinetics of raltegravir. Antimicrob Agents Chemother. 2009; 53: 2852–56.",
"DOI": null,
"article": 18
},
{
"id": 167,
"serial_number": 43,
"pmc": null,
"reference": "Song I, Borland J, Chen S, Lou Y, Peppercorn A, Wajima T et al. Effect of atazanavir and atazanavir/ritonavir on the pharmacokinetics of the next-generation HIV integrase inhibitor, S/GSK1349572. Br J Clin Pharmacol. 2011; 72: 103–08.",
"DOI": null,
"article": 18
},
{
"id": 168,
"serial_number": 44,
"pmc": null,
"reference": "Reese MJ, Savina PM, Generaux GT, Tracey H, Humphreys JE, Kanaoka E et al. In vitro investigations into the roles of drug transporters and metabolizing enzymes in the disposition and drug interactions of dolutegravir, a hiv integrase inhibitor. Drug Metab Dispos. 2013; 41: 353–61.",
"DOI": null,
"article": 18
},
{
"id": 169,
"serial_number": 45,
"pmc": null,
"reference": "Dooley KE, Sayre P, Borland J, Purdy E, Chen S, Song I et al. Safety, tolerability, and pharmacokinetics of the HIV integrase inhibitor dolutegravir given twice daily with rifampin or once daily with rifabutin: results of a phase 1 study among healthy subjects. J Acquir Immune Defic Syndr. 2013; 62: 21–27.",
"DOI": null,
"article": 18
},
{
"id": 170,
"serial_number": 46,
"pmc": null,
"reference": "Wenning LA, Friedman EJ, Kost JT, Breidinger SA, Stek JE, Lasseter KC et al. Lack of a significant drug interaction between raltegravir and tenofovir. Antimicrob Agents Chemother. 2008; 52: 3253–58.",
"DOI": null,
"article": 18
},
{
"id": 171,
"serial_number": 47,
"pmc": null,
"reference": "Iwamoto M, Wenning LA, Mistry GC, Petry AS, Liou SY, Ghosh K et al. Atazanavir modestly increases plasma levels of raltegravir in healthy subjects. Clin Infect Dis 2008; 47: 137–40.",
"DOI": null,
"article": 18
},
{
"id": 172,
"serial_number": 48,
"pmc": null,
"reference": "Weiss J, Rose J, Storch CH, Ketabi-Kiyanvash N, Sauer A, Haefeli WE et al. Modulation of human BCRP (ABCG2) activity by anti-HIV drugs. J Antimicrob Chemother. 2007; 59: 238–45.",
"DOI": null,
"article": 18
},
{
"id": 173,
"serial_number": 49,
"pmc": null,
"reference": "Hoque MT, Kis O, De Rosa MF, Bendayan R. Raltegravir permeability across blood-tissue barriers and the potential role of drug efflux transporters. Antimicrob Agents Chemother. 2015; 59: 2572–82.",
"DOI": null,
"article": 18
},
{
"id": 174,
"serial_number": 50,
"pmc": null,
"reference": "Hoetelmans RMW, Mariën K, De Pauw M, Hill A, Peeters M, Sekar V et al. Pharmacokinetic interaction between TMC114/ritonavir and tenofovir disoproxil fumarate in healthy volunteers. Br J Clin Pharmacol. 2007; 64: 655–61.",
"DOI": null,
"article": 18
},
{
"id": 175,
"serial_number": 51,
"pmc": null,
"reference": "Tourret J, Deray G, Isnard-Bagnis C. Tenofovir Effect on the Kidneys of HIV-Infected Patients: A Double-Edged Sword? J Am Soc Nephrol. 2013; 24: 1519–27.",
"DOI": null,
"article": 18
},
{
"id": 176,
"serial_number": 52,
"pmc": null,
"reference": "Perriëns JH, St Louis ME, Mukadi YB, Brown C, Prignot J, Pouthier F et al. Pulmonary tuberculosis in HIV-infected patients in Zaire. A controlled trial of treatment for either 6 or 12 months. N Engl J Med. 1995; 332: 779–84.",
"DOI": null,
"article": 18
},
{
"id": 177,
"serial_number": 53,
"pmc": null,
"reference": "McIlleron H, Meintjes G, Burman WJ, Maartens G. Complications of antiretroviral therapy in patients with tuberculosis: drug interactions, toxicity, and immune reconstitution inflammatory syndrome. J Infect Dis 2007; 196 Suppl: S63-75.",
"DOI": null,
"article": 18
},
{
"id": 178,
"serial_number": 54,
"pmc": null,
"reference": "Shipton LK, Wester CW, Stock S, Ndwapi N, Gaolathe T, Thior I et al. Safety and efficacy of nevirapine- and efavirenz-based antiretroviral treatment in adults treated for TB-HIV co-infection in Botswana. Int J Tuberc Lung Dis. 2009; 13: 360–66.",
"DOI": null,
"article": 18
},
{
"id": 179,
"serial_number": 55,
"pmc": null,
"reference": "La Porte CJL, Colbers EPH, Bertz R, Voncken DS, Wikstrom K, Boeree MJ et al. Pharmacokinetics of Adjusted-Dose Lopinavir-Ritonavir Combined with Rifampin in Healthy Volunteers. Antimicrob Agents Chemother. 2004; 48: 1553–60.",
"DOI": null,
"article": 18
},
{
"id": 180,
"serial_number": 56,
"pmc": null,
"reference": "Yimer G, Ueda N, Habtewold A, Amogne W, Suda A, Riedel KD et al. Pharmacogenetic & pharmacokinetic biomarker for efavirenz based ARV and rifampicin based anti-TB drug induced liver injury in TB-HIV infected patients. PLoS One 2011; 6.",
"DOI": null,
"article": 18
},
{
"id": 181,
"serial_number": 57,
"pmc": null,
"reference": "Nijland HMJ, L’homme RFA, Rongen GA, van Uden P, van Crevel R, Boeree MJ et al. High incidence of adverse events in healthy volunteers receiving rifampicin and adjusted doses of lopinavir/ritonavir tablets. AIDS. 2008; 22: 931–35.",
"DOI": null,
"article": 18
},
{
"id": 182,
"serial_number": 58,
"pmc": null,
"reference": "W HD, L KS, Laura L, A KM, Carol S, G GJ et al. Hepatotoxicity and Gastrointestinal Intolerance When Healthy Volunteers Taking Rifampin Add Twice-Daily Atazanavir and Ritonavir. JAIDS-JOURNAL Acquir IMMUNE Defic Syndr 2009; 50: 290–93.",
"DOI": null,
"article": 18
},
{
"id": 183,
"serial_number": 59,
"pmc": null,
"reference": "Schmitt C, Riek M, Winters K, Schutz M, Grange S. Unexpected hepatotoxicity of rifampin and saquinavir/ritonavir in healthy male volunteers. Arch Drug Inf. 2009; 2: 8–16.",
"DOI": null,
"article": 18
},
{
"id": 184,
"serial_number": 60,
"pmc": null,
"reference": "Benet LZ. The drug transporter-metabolism alliance: Uncovering and defining the interplay. Mol. Pharm. 2009; 6: 1631–43.",
"DOI": null,
"article": 18
},
{
"id": 185,
"serial_number": 61,
"pmc": null,
"reference": "Karlgren M, Ahlin G, Bergström CAS, Svensson R, Palm J, Artursson P. In Vitro and in silico strategies to identify OATP1B1 Inhibitors and Predict Clinical Drug-Drug Interactions. Pharm Res. 2012; 29: 411–26.",
"DOI": null,
"article": 18
},
{
"id": 186,
"serial_number": 62,
"pmc": null,
"reference": "Benet LZ, Cummins CL. The drug efflux-metabolism alliance: Biochemical aspects. In: Advanced Drug Delivery Reviews. 2001 doi:10.1016/S0169-409X(01)00178-8.",
"DOI": null,
"article": 18
},
{
"id": 187,
"serial_number": 63,
"pmc": null,
"reference": "Wacher VJ, Wu CY, Benet LZ. Overlapping substrate specificities and tissue distribution of cytochrome p450 3A and P-glycoprotein: Implications for drug delivery and activity in cancer chemotherapy. Mol. Carcinog. 1995; 13: 129–34.",
"DOI": null,
"article": 18
},
{
"id": 188,
"serial_number": 64,
"pmc": null,
"reference": "Zhang Y, Benet LZ. The gut as a barrier to drug absorption: combined role of cytochrome P450 3A and P-glycoprotein. Clin Pharmacokinet. 2001; 40: 159–68.",
"DOI": null,
"article": 18
},
{
"id": 189,
"serial_number": 65,
"pmc": null,
"reference": "Lown KS, Mayo RR, Leichtman AB, Hsiao HL, Turgeon DK, Schmiedlin-Ren P et al. Role of intestinal P-glycoprotein (mdr1) in interpatient variation in the oral bioavailability of cyclosporine. Clin Pharmacol Ther. 1997; 62: 248–60.",
"DOI": null,
"article": 18
}
]
},
{
"id": 17,
"slug": "178-1513541446-choice-of-samples-in-extracellular-microrna-research-which-fraction-is-better-exosomal-or-nonexosomal",
"featured": false,
"slider": false,
"issue": "Vol1 Issue1",
"type": "review_article",
"manuscript_id": "178-1513541446",
"recieved": "2017-12-17",
"revised": null,
"accepted": "2017-12-31",
"published": "2018-01-15",
"pdf_file": "https://jabet.bsmiab.org/media/pdf_file/2023/47/178-1513541446.pdf",
"title": "Choice of samples in extracellular microRNA research: Which fraction is better- exosomal or nonexosomal?",
"abstract": "<p>MicroRNAs (miRNAs) are a species of noncoding RNA that post-transcriptionally regulates the expression of target mRNAs via degradation or translational repression. Recently, miRNAs in biofluids got extraordinary research priority in the field of noninvasive biomarker development because of their association with pathophysiological events, noninvasive way to obtain, and are markable stability in the extracellular harsh environment. Although several hypotheses have been proposed, the precise mechanisms of cellular release of extracellular miRNAs are not properly understood. In addition, it is often confusing to the researchers, which fraction of biofluid (based on release mechanisms) is suitable for biomarker development research. This review will briefly introduce extracellular miRNAs (EC miRNAs) and their release mechanisms. Furthermore, based on recent evidence, it will also summarize and suggest the appropriate fraction of biofluid to work with for biomarker development.</p>",
"journal_reference": "J Adv Biotechnol Exp Ther. 2018; 1(1) : 11-16.",
"academic_editor": "Md Nurul Haider, PhD; Bangladesh Agricultural University, Bangladesh",
"cite_info": "Sohel MMH. Choice of samples in extracellular microRNA research: Which fraction is better- exosomal or nonexosomal? J Adv Biotechnol Exp Ther. 2018; 1(1) : 11-16.",
"keywords": [
"Circulating microRNA",
"nonexosomal",
"extracellular microRNA",
"exosomal",
"serum/plasma"
],
"DOI": "10.5455/jabet.d3",
"sections": [
{
"section_number": 1,
"section_title": "microRNAs AND EXTRACELLULAR microRNAs",
"body": "<p>microRNAs (miRNAs) are endogenously initiated noncoding RNA species of 18-24 nucleotide that post-transcriptionally regulate the expression of target mRNAs either via mRNA destabilization or transcriptional repression [<a href=\"#r-1\">1</a>]. It is estimated that miRNAs cover 1-5% of the animal genome and bioinformatic studies predicted that almost 60% mammalian genes can be targeted by one or more miRNAs [<a href=\"#r-2\">2</a>], thus they play a central role in cellular proliferation, migration, differentiation, survival, and cell fate. Since their discovery in the early 1990s [<a href=\"#r-3\">3</a>], miRNAs have been broadly studied in different organism ranging from single- to multi-cellular organisms and found to be involved in important cellular pathways. According to the miRbase 21, more than 28000 miRNAs have been reported in all organisms that could potentially influence the expression of genes and associated with pathophysiological conditions [<a href=\"#r-4\">4</a>].<br />\r\nWhile the majority of the miRNAs are detected intracellularly, a handful of miRNAs are found in various body fluids and commonly known as circulating miRNA or circulatory miRNA or extracellular miRNAs (ECmiRNAs). The first report of ECmiRNAs in biofluid was made by Chim and his colleagues [<a href=\"#r-5\">5</a>] where they observed that placental miRNAs are readily detectable in maternal blood plasma. At the same time, another report claimed the existence of tumor-associated ECmiRNAs in the serum sample of the patients with diffused B-cell lymphoma [<a href=\"#r-6\">6</a>]. The discovery of ECmiRNAs in blood serum/plasma opened a new era of miRNA research and a series of studies have consistently reported that the presence of ECmiRNAs in other biofluids including plasma [<a href=\"#r-7\">7</a>], follicular fluid [<a href=\"#r-8\">8</a>,<a href=\"#r-9\">9</a>], amniotic fluid, and almost every other biofluids [<a href=\"#r-10\">10</a>]. However, the total miRNA concentration, number of detected miRNAs, and level of detection vary significantly among different fluid types [<a href=\"#r-10\">10</a>]. In addition, the expression profile of ECmiRNAs in certain biofluids in relation to different pathophysiological conditions indicates that ECmiRNAs could be selectively released from healthy cells [<a href=\"#r-11\">11</a>–<a href=\"#r-13\">13</a>] and has a great potential to serve as biomarkers to assess the pathological conditions.</p>"
},
{
"section_number": 2,
"section_title": "CHARACTERISTICS OF ECmiRNAs",
"body": "<p>ECmiRNAs have distinct characteristics which are completely absent in cellular miRNAs. For instance, the introduction of cellular or synthetic miRNAs in plasma or any other fluid results in rapid degradation (within few minutes) and the addition of denaturing solution protect exogenous miRNAs from degradation by inactivating RNase activity in plasma [<a href=\"#r-14\">14</a>]. In contrast, ECmiRNAs can stay for a long time in circulation or other biofluids without degradation and exhibit a higher level of resistance against the activity of high endogenous RNase indicating that ECmiRNAs avoid RNase activity by wrapping themselves with some protective manner [<a href=\"#r-15\">15</a>]. While cellular miRNAs are susceptible to the unusual conditions like low or high pH, boiling, and prolonged storage time, ECmiRNAs showed remarkable stability under such conditions. In addition, ECmiRNAs are detectable in qRT-PCR after multiple freeze-thaw cycles (even up to 10 cycles) whereas cellular miRNAs start their signals after few freeze-thaw cycles [<a href=\"#r-14\">14</a>,<a href=\"#r-16\">16</a>]. In addition, Mitchell and his colleagues reported that a 24 h incubation of ECmiRNAs derived from serum at room temperature could not affect the specific expression patterns in qPCR [<a href=\"#r-17\">17</a>]. In addition, Chen and colleagues reported that ECmiRNAs isolated from serum showed resistance against the treatment of RNase A, compared to other endogenous RNAs [<a href=\"#r-15\">15</a>]. Majority of the serum derived ECmiRNAs are considerably expressed after being subjected to 3 h or overnight RNase treatment, on the other hand, large RNA species including 18s/28s rRNA, β-actin, GAPDH, and U6 degraded and showed no expression [<a href=\"#r-15\">15</a>].<br />\r\nAlthough the release mechanism and remarkable stability of ECmiRNAs in bio-fluids are poorly understood, several hypotheses have been proposed to explain the mechanism. Nucleoprotein bound existence of RNAs is one of the earliest theory where the DNA fraction of the complex binds with RNA to form a DNA-RNA heterodimer [<a href=\"#r-18\">18</a>]. Unexpectedly, a couple of years later, it has been reported that the protection and existence of RNA molecules in plasma or serum probably due to the binding with lipoprotein complex or lipid molecules, not with the DNA fraction of nucleoprotein [<a href=\"#r-19\">19</a>].</p>"
},
{
"section_number": 3,
"section_title": "MODE OF RELEASE OF ECmiRNAs",
"body": "<p>The mechanism of remarkable stability of ECmiRNAs in biofluids provoked us to speculate that these miRNAs are encapsulated by some manners to avoid degradation. Interestingly, exosome-mediated release, existence, and transport of miRNAs and mRNAs in cultured cells have reported shortly before the discovery of ECmiRNAs [<a href=\"#r-20\">20</a>] which strongly suggests the hypothesis of the release of ECmiRNAs through extracellular vesicles. The membranes of extracellular vesicles are impermeable to RNase and this can keep the encapsulated miRNAs safe from degradation in the presence of high RNase activity in serum/plasma. Therefore, vesicles encapsulated release and existence of miRNAs in extracellular environment explained elegantly the remarkable stability of ECmiRNAs [<a href=\"#r-21\">21</a>]. Later, Hunter and his colleagues reported that purified human peripheral blood microvesicles contain miRNAs [<a href=\"#r-22\">22</a>] which further reinforce the theory of microvesicles encapsulated miRNAs in the extracellular environment.<br />\r\nExtracellular vesicles are a combination of a heterogeneous population of exosomes and microparticles (shedding vesicles). Exosomes are homologous small vesicles (50-90 nm) [<a href=\"#r-23\">23</a>], whereas microparticles/microvesicles (MVs) are lipid vesicles that are <1 μm in diameter and are secreted into extracellular environment by different types of cells and platelets [<a href=\"#r-24\">24</a>]. While exosomes are generated via endocytic recycling pathway (inward budding), MVs are produced from plasma membrane via outward budding and fission. Thus, the membrane composition of MVs is much similar to the plasma membrane of their parent cells. Hence the membrane composition of MVs is predominantly depended on the cell of origin [<a href=\"#r-25\">25</a>]. In the following years, several studies reported the existence of miRNAs in various biofluid in association with either exosomes [<a href=\"#r-12\">12</a>,<a href=\"#r-23\">23</a>] or microvesicles [<a href=\"#r-26\">26</a>,<a href=\"#r-27\">27</a>].<br />\r\nIn 2011, the vesicle-enwrapped release and existence of miRNAs were hugely challenged when independent two groups claimed that a large fraction of ECmiRNAs exists in the cell culture media [14] and in human plasma [<a href=\"#r-28\">28</a>] that are outside of shedding vesicles and exosomes. In addition, authors claimed that only 10% of ECmiRNAs are released in plasma through vesicles and potentially 90% of them are associated with protein complexes [<a href=\"#r-28\">28</a>]. Furthermore, when size-exclusion chromatography was used to complete separation of vesicles from protein complex, it showed that most of the ECmiRNAs are ribonucleoprotein complex associated and only a few miRNAs are predominantly associated with microvesicles [<a href=\"#r-28\">28</a>]. It is important to note that all mature miRNAs become associated with the AGO protein families during their intracellular synthesis process [<a href=\"#r-2\">2</a>]. Therefore, it is logical to think that ECmiRNAs could exist with AGO protein, particularly AGO2 protein, in the extracellular environment. In addition, it has been known for a long time that AGO2 proteins are remarkably stable in nuclease- and protein-rich environment which also explained the stability of protein complex associated ECmiRNAs [<a href=\"#r-14\">14</a>]. Later, Gallo and colleagues reported that several miRNAs including miR-16 and miR-92a were detected in the pellet of ultracentrifuges serum samples while they are barely detected in the supernatant [<a href=\"#r-29\">29</a>]. It was unexpected because Arroyo and colleagues reported that miR-16 and miR-92a were predominantly associated with nucleoprotein [<a href=\"#r-28\">28</a>]. The difference in the results of two groups may be due to the differences in the sample processing, isolation, and data normalization method.</p>\r\n\r\n<p>In the following years of ECmiRNA discovery, accumulated studies have reported that ECmiRNAs are packaged into apoptotic bodies[<a href=\"#r-30\">30</a>], high-density lipoprotein (HDL) particles [<a href=\"#r-31\">31</a>], microvesicles [<a href=\"#r-22\">22</a>], exosomes [<a href=\"#r-23\">23</a>], and AGO2 proteins [<a href=\"#r-28\">28</a>]. A pictorial representation of release and existence of extracellular miRNAs in biofluids are presented in <a href=\"#figure1\">Figure 1</a>. The inclusion of apoptotic body associated miRNAs in ECmiRNA category is questionable due to its size (comparable with the size of cellular debris and platelets). In addition, in the commonly used protocol for ECmiRNA isolation, during the sample processing step, apoptotic bodies are removed with cell debris in the initial ultracentrifugation step. Thus, this fraction of ECmiRNAs is overlooked and understudied.</p>\r\n\r\n<div id=\"figure1\">\r\n<figure class=\"image\"><img alt=\"\" height=\"357\" src=\"/media/article_images/2023/38/29/178-1513541446-Figure1.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 1: </strong>Biogenesis and extracellular release of miRNAs. Mature miRNAs bind with protein complex (primarily AGO2) either to form RISC complex or released in the extracellular environment. They can also be loaded to exosomes and released. Similarly, EC miRNAs can be encapsulated via microvesicles or bind with HDL and released from cells to the extracellular environment. This figure is adapted from [<a href=\"#r-4\">4</a>].</figcaption>\r\n</figure>\r\n</div>"
},
{
"section_number": 4,
"section_title": "CHOICE OF SAMPLES FOR ECmiRNA RESEARCH",
"body": "<p>Soon after the discovery of ECmiRNAs, they received an utmost research priority due to their potential as a noninvasive biomarker and to understand the disease pathogenesis. Therefore, much of the efforts in profiling ECmiRNAs in various biofluid is involved in investigating the expression patterns of ECmiRNAs and correlates them with different physiological status and disease progression. Despite the widespread interests in developing biomarkers based on ECmiRNAs, heterogeneity among the results in different studies have been observed and hence the use of ECmiRNAs as reliable and reproducible biomarker remain in infancy [<a href=\"#r-4\">4</a>]. The contributing factors in the variation in results can be both preanalytical and analytical. Preanalytical factors are starting material, methods of collection, processing, RNA extraction, and expression analysis. The source and composition of these biofluids are quite different; therefore, the processing and isolation methods need to be changed/adopted according to the origin of biofluids. Thus, to avoid the variation in the results of ECmiRNA research, a standardized protocol should be followed from sample collection to data analysis [<a href=\"#r-4\">4</a>].<br />\r\nOne of the major points to be considered in ECmiRNA research is the choice of sample or starting materials. For instance, when ECmiRNAs are quantified from circulation, a critical decision should be made that is which fraction will be the starting material- whole blood or sera or plasma? Essentially, we must discard whole blood from the list as it contains cellular fractions (white and red blood cells) [<a href=\"#r-32\">32</a>]. The basic difference between serum and plasma is the absence or presence of clotting factors, respectively. Both plasma and serum are used for ECmiRNA research. If plasma is the starting choice, ethylenediaminetetraacetic acid (EDTA) is the better choice over heparin as it is known as PCR inhibitor. Although many studies investigated plasma and serum side-by-side and found no or little difference in ECmiRNA quantification [<a href=\"#r-15\">15</a>], consistently higher concentrations were found in serum samples [<a href=\"#r-33\">33</a>]. Similarly, cellular contribution in other biofluids should be discarded before subjected to ECmiRNA expression analysis. This could be achieved by using a centrifugation step at 25,000 × g for 30 min [<a href=\"#r-12\">12</a>]. In addition to cellular fraction, the centrifugation step removes the cellular debris, apoptotic bubbles, and fractions of large macroparticles.<br />\r\nNow, in the cell-free centrifuged biofluid, ECmiRNAs can be existed by two means such as exosomal and nonexosomal (includes microvesicles, AGO2 protein, and HDL).When AGO2 protein-coupled ECmiRNAs are kept for several weeks in cell lysates without protease inhibitors, it showed no degradation. This clearly suggests that the protein complex associated ECmiRNAs may simply derive from the passive release of cytosolic components during cell death [<a href=\"#r-14\">14</a>]. HDL and LDL can carry a pure population of ECmiRNA in biofluids [<a href=\"#r-31\">31</a>] and their functional relevance has been demonstrated in themodel organism [<a href=\"#r-34\">34</a>] and human [<a href=\"#r-35\">35</a>]. However, it is not entirely clear that HDL coupled ECmiRNAs are selectively and actively released from the donor cells and spontaneously taken up by the recipient cells. Therefore, it is clear that nonexosomal ECmiRNAs are the result of nonspecific and passive release from the cells and may not be the better choice for the biomarker study.<br />\r\nExosome formation and release is a tightly coordinated and complex process which is orchestrated by enzymatic activation and energy in the form of adenosine triphosphate (ATP) [<a href=\"#r-36\">36</a>], and the expression patterns of exosomal ECmiRNAs(Exo-miRs)obtained from culture media and various bio-fluids derived from bio-fluids are different from their cell of origin suggesting there might be an active selection mechanism for exosome release and their contents. In contrast to nonexosomal ECmiRNAs, exosomal ECmiRNAs (Exo-miRs) are released from viable cells and can be taken up by the recipient cells and modulate the expression of genes [<a href=\"#r-12\">12</a>] and trigger functional effects [<a href=\"#r-20\">20</a>,<a href=\"#r-37\">37</a>], however, the sorting and secretion of Exo-miRs are not fully understood. Pegtel and colleagues demonstrated the potential of Exo-miRs to facilitate viral infection [<a href=\"#r-37\">37</a>] while Sohel and colleagues showed the gene expression modulation capacity of follicular fluid derived Exo-miRs in the recipient cells [<a href=\"#r-12\">12</a>]. Many other studies have further indicated that miRNAs can be transported to the recipient cells via exosomes and could facilitate the cell-to-cell communication <em>in vitro </em>[<a href=\"#r-20\">20</a>,<a href=\"#r-38\">38</a>] and <em>in vivo </em>[<a href=\"#r-39\">39</a>]. Exo-miRs are particularly important in cancer biomarker development. The main reason behind this is- cancer cells, in a given microenvironment, constitutively secrete exosomes to mediate cell-cell communication with same neighboring cells (autocrine manner), with different neighboring cells (paracrine manner), and with distant cells (endocrine manner) [<a href=\"#r-40\">40</a>]. Therefore, much of the efforts were made in developing cancer biomarkers using Exo-miRs whereas nonexosomal ECmiRNAs were overlooked.<br />\r\nECmiRNAs are the new celebrity of miRNA research where Exo-miRs dominating the field. The specific expression pattern of Exo-miRs in various biofluids is distinctly associated with various pathophysiological conditions. Thus, Exo-miRs have the great potential in biomarker development and perhaps, in future, Exo-miRs will be routinely used for therapeutic purpose. Indeed, among all ECmiRNAs, Exo-miRs are released from viable cells and have the ability to modulate the biological function in recipient cells, thus, the exosomal fraction of biofluid is always beena better choice over nonexosomal fraction for biomarker development and/or therapeutic study.</p>"
},
{
"section_number": 5,
"section_title": "CONFLICT OF INTEREST",
"body": "<p>The author declares that no conflict of interest exists.</p>"
},
{
"section_number": 6,
"section_title": "FUNDING",
"body": "<p>This research does not receive any fund from public or privet organization.</p>"
}
],
"figures": [
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/38/29/178-1513541446-Figure1.jpg",
"caption": "Figure 1. Biogenesis and extracellular release of miRNAs. Mature miRNAs bind with protein complex (primarily AGO2) either to form RISC complex or released in the extracellular environment. They can also be loaded to exosomes and released. Similarly, EC miRNAs can be encapsulated via microvesicles or bind with HDL and released from cells to the extracellular environment. This figure is adapted from [4].",
"featured": true
}
],
"authors": [
{
"id": 28,
"affiliation": [
{
"affiliation": "Department of Animal Science, Faculty of Agriculture, Erciyes University, Kayseri 38039, Turkey"
},
{
"affiliation": "Genome and Stem Cell Centre, Erciyes University, Kayseri 38039, Turkey."
}
],
"first_name": "Md Mahmodul Hasan",
"family_name": "Sohel",
"email": "sohel.mmh@gmail.com",
"author_order": 1,
"ORCID": "https://orcid.org/0000-0003-2224-085X",
"corresponding": true,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "Md Mahmodul Hasan Sohel, Department of Animal Science, Faculty of Agriculture, Erciyes University, Kayseri 38039, Turkey, e-mail: sohel@erciyes.edu.tr, or sohel.mmh@gmail.com",
"article": 17
}
],
"views": 695,
"downloads": 82,
"references": [
{
"id": 85,
"serial_number": 1,
"pmc": null,
"reference": "Tesfaye D, Salilew-Wondim D, Gebremedhn S, Sohel MMH, Pandey HO, Hoelker M et al. Potential role of microRNAs in mammalian female fertility. Reprod Fertil Dev. 2017; 29: 8–23.",
"DOI": null,
"article": 17
},
{
"id": 86,
"serial_number": 2,
"pmc": null,
"reference": "Hossain MM, Sohel MMH, Schellander K, Tesfaye D. Characterization and importance of microRNAs in mammalian gonadal functions. Cell Tissue Res. 2012; 349: 679–90.",
"DOI": null,
"article": 17
},
{
"id": 87,
"serial_number": 3,
"pmc": null,
"reference": "Lee RC, Feinbaum RL, Ambros V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993; 75: 843–54.",
"DOI": null,
"article": 17
},
{
"id": 88,
"serial_number": 4,
"pmc": null,
"reference": "Sohel MMH. Extracellular/Circulating MicroRNAs: Release Mechanisms, Functions and Challenges. Achiev Life Sci 2016; 10: 175–186.",
"DOI": null,
"article": 17
},
{
"id": 89,
"serial_number": 5,
"pmc": null,
"reference": "Chim SSC, Shing TKF, Hung ECW, Leung T-Y, Lau T-K, Chiu RWK et al. Detection and characterization of placental microRNAs in maternal plasma. Clin Chem 2008; 54: 482–90.",
"DOI": null,
"article": 17
},
{
"id": 90,
"serial_number": 6,
"pmc": null,
"reference": "Lawrie CH, Gal S, Dunlop HM, Pushkaran B, Liggins AP, Pulford K et al. Detection of elevated levels of tumour-associated microRNAs in serum of patients with diffuse large B-cell lymphoma. Br J Haematol 2008; 141: 672–675.",
"DOI": null,
"article": 17
},
{
"id": 91,
"serial_number": 7,
"pmc": null,
"reference": "Noferesti SS, Hoelker M, Sohel MMH, Salilew-Wondim D, Rings F, Schellander K et al. Effect of ovarian hyperstimulation on circulatory microRNA profile in bovine follicular fluid and blood plasma. Reprod Fertil Dev 2012; 25: 251.",
"DOI": null,
"article": 17
},
{
"id": 92,
"serial_number": 8,
"pmc": null,
"reference": "Sohel MMH, Salilew-Wondim D, Hölker M, Rings F, Schellander K, Tesfaye D. Circulatory microRNA signatures in follicular fluid in relation to the growth status of bovine oocytes. Reprod Fertil Dev 2012; 25: 251.",
"DOI": null,
"article": 17
},
{
"id": 93,
"serial_number": 9,
"pmc": null,
"reference": "Sohel MMH, Noferesti SS, Salilew-Wondim D, Hoelker M, Rings F, Tholen E et al. Relative abundance of extra-cellular miRNAs in bovine follicular fluid: Implication for cell–cell communication during oocyte growth. Anim Reprod Sci 2014; 149: 98.",
"DOI": null,
"article": 17
},
{
"id": 94,
"serial_number": 10,
"pmc": null,
"reference": "Weber JA, Baxter DH, Zhang S, Huang DY, Huang KH, Lee MJ et al. The microRNA spectrum in 12 body fluids. Clin Chem 2010; 56: 1733–41.",
"DOI": null,
"article": 17
},
{
"id": 95,
"serial_number": 11,
"pmc": null,
"reference": "Pigati L, Yaddanapudi SCS, Iyengar R, Kim D-J, Hearn SA, Danforth D et al. Selective release of microRNA species from normal and malignant mammary epithelial cells. PLoS One 2010; 5: e13515.",
"DOI": null,
"article": 17
},
{
"id": 96,
"serial_number": 12,
"pmc": null,
"reference": "Sohel MMH, Hoelker M, Noferesti SS, Salilew-Wondim D, Tholen E, Looft C et al. Exosomal and non-exosomal transport of extra-cellular microRNAs in follicular fluid: Implications for bovine oocyte developmental competence. PLoS One 2013; 8. doi:10.1371/journal.pone.0078505.",
"DOI": null,
"article": 17
},
{
"id": 97,
"serial_number": 13,
"pmc": null,
"reference": "Noferesti SS, Sohel MMH, Hoelker M, Salilew-Wondim D, Tholen E, Looft C et al. Controlled ovarian hyperstimulation induced changes in the expression of circulatory miRNA in bovine follicular fluid and blood plasma. J Ovarian Res 2015; 8: 81.",
"DOI": null,
"article": 17
},
{
"id": 98,
"serial_number": 14,
"pmc": null,
"reference": "Turchinovich A, Weiz L, Langheinz A, Burwinkel B. Characterization of extracellular circulating microRNA. Nucleic Acids Res 2011; 39: 7223–33.",
"DOI": null,
"article": 17
},
{
"id": 99,
"serial_number": 15,
"pmc": null,
"reference": "Chen X, Ba Y, Ma L, Cai X, Yin Y, Wang K et al. Characterization of microRNAs in serum: a novel class of biomarkers for diagnosis of cancer and other diseases. Cell Res 2008; 18: 997–1006.",
"DOI": null,
"article": 17
},
{
"id": 100,
"serial_number": 16,
"pmc": null,
"reference": "Taylor DD, Gercel-Taylor C. MicroRNA signatures of tumor-derived exosomes as diagnostic biomarkers of ovarian cancer. Gynecol Oncol 2008; 110: 13–21.",
"DOI": null,
"article": 17
},
{
"id": 101,
"serial_number": 17,
"pmc": null,
"reference": "Mitchell PS, Parkin RK, Kroh EM, Fritz BR, Wyman SK, Pogosova-Agadjanyan EL et al. Circulating microRNAs as stable blood-based markers for cancer detection. Proc Natl Acad Sci U S A 2008; 105: 10513–8.",
"DOI": null,
"article": 17
},
{
"id": 102,
"serial_number": 18,
"pmc": null,
"reference": "Sisco KL. Is RNA in serum bound to nucleoprotein complexes? Clin Chem 2001; 47: 1744–5.",
"DOI": null,
"article": 17
},
{
"id": 103,
"serial_number": 19,
"pmc": null,
"reference": "El-Hefnawy T, Raja S, Kelly L, Bigbee WL, Kirkwood JM, Luketich JD et al. Characterization of amplifiable, circulating RNA in plasma and its potential as a tool for cancer diagnostics. Clin Chem 2004; 50: 564–73.",
"DOI": null,
"article": 17
},
{
"id": 104,
"serial_number": 20,
"pmc": null,
"reference": "Valadi H, Ekström K, Bossios A, Sjöstrand M, Lee JJ, Lötvall JO. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat Cell Biol 2007; 9: 654–9.",
"DOI": null,
"article": 17
},
{
"id": 105,
"serial_number": 21,
"pmc": null,
"reference": "Turchinovich A, Weiz L, Burwinkel B. Extracellular miRNAs: the mystery of their origin and function. Trends Biochem Sci 2012; 37: 460–5.",
"DOI": null,
"article": 17
},
{
"id": 106,
"serial_number": 22,
"pmc": null,
"reference": "Hunter MP, Ismail N, Zhang X, Aguda BD, Lee EJ, Yu L et al. Detection of microRNA expression in human peripheral blood microvesicles. PLoS One 2008; 3: e3694.",
"DOI": null,
"article": 17
},
{
"id": 107,
"serial_number": 23,
"pmc": null,
"reference": "Camussi G, Deregibus MC, Bruno S, Cantaluppi V, Biancone L. Exosomes/microvesicles as a mechanism of cell-to-cell communication. Kidney Int 2010; 78: 838–48.",
"DOI": null,
"article": 17
},
{
"id": 108,
"serial_number": 24,
"pmc": null,
"reference": "Aharon A, Katzenell S, Tamari T, Brenner B. Microparticles bearing tissue factor and tissue factor pathway inhibitor in gestational vascular complications. J Thromb Haemost 2009; 7: 1047–50.",
"DOI": null,
"article": 17
},
{
"id": 109,
"serial_number": 25,
"pmc": null,
"reference": "Lee Y, El Andaloussi S, Wood MJA. Exosomes and microvesicles: extracellular vesicles for genetic information transfer and gene therapy. Hum Mol Genet 2012; 21: R125-34.",
"DOI": null,
"article": 17
},
{
"id": 110,
"serial_number": 26,
"pmc": null,
"reference": "Yuan A, Farber EL, Rapoport AL, Tejada D, Deniskin R, Akhmedov NB et al. Transfer of microRNAs by embryonic stem cell microvesicles. PLoS One 2009; 4: e4722.",
"DOI": null,
"article": 17
},
{
"id": 111,
"serial_number": 27,
"pmc": null,
"reference": "Castellana D, Kunzelmann C, Freyssinet J-M. Pathophysiologic significance of procoagulant microvesicles in cancer disease and progression. Hämostaseologie 2009; 29: 51–7.",
"DOI": null,
"article": 17
},
{
"id": 112,
"serial_number": 28,
"pmc": null,
"reference": "Arroyo JD, Chevillet JR, Kroh EM, Ruf IK, Pritchard CC, Gibson DF et al. Argonaute2 complexes carry a population of circulating microRNAs independent of vesicles in human plasma. Proc Natl Acad Sci U S A 2011; 108: 5003–8.",
"DOI": null,
"article": 17
},
{
"id": 113,
"serial_number": 29,
"pmc": null,
"reference": "Gallo A, Tandon M, Alevizos I, Illei GG. The majority of microRNAs detectable in serum and saliva is concentrated in exosomes. PLoS One 2012; 7: e30679.",
"DOI": null,
"article": 17
},
{
"id": 114,
"serial_number": 30,
"pmc": null,
"reference": "Zernecke A, Bidzhekov K, Noels H, Shagdarsuren E, Gan L, Denecke B et al. Delivery of microRNA-126 by apoptotic bodies induces CXCL12-dependent vascular protection. Sci Signal 2009; 2: ra81.",
"DOI": null,
"article": 17
},
{
"id": 115,
"serial_number": 31,
"pmc": null,
"reference": "Vickers KC, Palmisano BT, Shoucri BM, Shamburek RD, Remaley AT. MicroRNAs are transported in plasma and delivered to recipient cells by high-density lipoproteins. Nat Cell Biol 2011; 13: 423–433.",
"DOI": null,
"article": 17
},
{
"id": 116,
"serial_number": 32,
"pmc": null,
"reference": "Pritchard CC, Kroh E, Wood B, Arroyo JD, Dougherty KJ, Miyaji MM et al. Blood cell origin of circulating microRNAs: a cautionary note for cancer biomarker studies. Cancer Prev Res (Phila) 2012; 5: 492–7.",
"DOI": null,
"article": 17
},
{
"id": 117,
"serial_number": 33,
"pmc": null,
"reference": "Wang K, Yuan Y, Cho J-H, McClarty S, Baxter D, Galas DJ. Comparing the MicroRNA spectrum between serum and plasma. PLoS One 2012; 7: e41561.",
"DOI": null,
"article": 17
},
{
"id": 118,
"serial_number": 34,
"pmc": null,
"reference": "Tabet F, Vickers KC, Cuesta Torres LF, Wiese CB, Shoucri BM, Lambert G et al. HDL-transferred microRNA-223 regulates ICAM-1 expression in endothelial cells. Nat Commun 2014; 5: 3292.",
"DOI": null,
"article": 17
},
{
"id": 119,
"serial_number": 35,
"pmc": null,
"reference": "Niculescu LS, Simionescu N, Sanda GM, Carnuta MG, Stancu CS, Popescu AC et al. MiR-486 and miR-92a Identified in Circulating HDL Discriminate between Stable and Vulnerable Coronary Artery Disease Patients. PLoS One 2015; 10: e0140958.",
"DOI": null,
"article": 17
},
{
"id": 120,
"serial_number": 36,
"pmc": null,
"reference": "Yáñez-Mó M, Siljander PR-M, Andreu Z, Zavec AB, Borràs FE, Buzas EI et al. Biological properties of extracellular vesicles and their physiological functions. J Extracell vesicles 2015; 4: 27066.",
"DOI": null,
"article": 17
},
{
"id": 121,
"serial_number": 37,
"pmc": null,
"reference": "Pegtel DM, Cosmopoulos K, Thorley-Lawson DA, van Eijndhoven MAJ, Hopmans ES, Lindenberg JL et al. Functional delivery of viral miRNAs via exosomes. Proc Natl Acad Sci U S A 2010; 107: 6328–33.",
"DOI": null,
"article": 17
},
{
"id": 122,
"serial_number": 38,
"pmc": null,
"reference": "Montecalvo A, Larregina AT, Shufesky WJ, Stolz DB, Sullivan MLG, Karlsson JM et al. Mechanism of transfer of functional microRNAs between mouse dendritic cells via exosomes. Blood 2012; 119: 756–66.",
"DOI": null,
"article": 17
},
{
"id": 123,
"serial_number": 39,
"pmc": null,
"reference": "da Silveira JC, Veeramachaneni DNR, Winger QA, Carnevale EM, Bouma GJ. Cell-Secreted Vesicles in Equine Ovarian Follicular Fluid Contain miRNAs and Proteins: A Possible New Form of Cell Communication Within the Ovarian Follicle. Biol Reprod 2012; 86: 71–71.",
"DOI": null,
"article": 17
},
{
"id": 124,
"serial_number": 40,
"pmc": null,
"reference": "Tomasetti M, Lee W, Santarelli L, Neuzil J. Exosome-derived microRNAs in cancer metabolism: possible implications in cancer diagnostics and therapy. Exp Mol Med 2017; 49: e285–e285.",
"DOI": null,
"article": 17
}
]
},
{
"id": 19,
"slug": "178-1513778843-a-protein-isolation-method-for-western-blot-to-study-histones-with-an-internal-control-protein",
"featured": false,
"slider": false,
"issue": "Vol1 Issue1",
"type": "short_communication",
"manuscript_id": "178-1513778843",
"recieved": "2017-09-15",
"revised": null,
"accepted": "2017-12-21",
"published": "2018-01-05",
"pdf_file": "https://jabet.bsmiab.org/media/pdf_file/2023/08/178-1513778843.pdf",
"title": "A protein isolation method for western blot to study histones with an internal control protein",
"abstract": "<p>Histone modification is one of the attractive targets for epigenetic studies. However, current methods to extract chromatin-associated proteins for western blot of histone modification have some weaknesses such as the loss of housekeeping proteins. In this study, we are presenting a simple method to isolate nuclear protein for studying histone modification by immunoblotting with housekeeping proteins. This method provided high protein concentration from minute tissue samples and importantly, it allowed us to detect acetylated histones together with internal control proteins such as β actin.</p>",
"journal_reference": "J Adv Biotechnol Exp Ther. 2018; 1(1) : 25-28",
"academic_editor": "Dr. Md Mahmodul Hasan Sohel, Erciyes University, Turkey.",
"cite_info": "Chu DT, Tao Y. A protein isolation method for western blot to study histones with an internal control protein. J Adv Biotechnol Exp Ther. 2018; 1(1) : 25-28.",
"keywords": [
"Protein isolation",
"housekeeping protein",
"histone modification"
],
"DOI": "10.5455/jabet.d4",
"sections": [
{
"section_number": 1,
"section_title": "SHORT COMMUNICATION",
"body": "<p>Epigenetics has been emerging as an attractive field in biomedical sciences. Somatic changes within an individual or between individuals, which are not caused by changes in DNA sequence such as mutation, are promising targets for epigenetic studies, where DNA methylation and histone modifications are mostly considered. DNA methylation, one of the most important mechanisms controlling the gene expression, is established early in development [<a href=\"#r-1\">1</a>]. Histone modifications regulating the manipulation and expression of DNA in most biological processes [<a href=\"#r-2\">2</a>] which may occur throughout the life under different conditions.</p>\r\n\r\n<p>To study the epigenetic regulation of gene expression, we have some good techniques such as western blot, DNA methylation assay, and chromatin immunoprecipitation (ChiP) assay. Western blot seems to be method considered as a simple and cheap technique to investigate global histone modifications applied in several epigenetic studies [<a href=\"#r-3\">3</a>-<a href=\"#r-4\">4</a>]. However, the traditional methods of nuclear protein isolation and histone extraction for immunoblotting of global histone modifications have some weaknesses including the low yield of isolated proteins and do not allow to have internal controls [<a href=\"#r-3\">3</a>, <a href=\"#r-5\">5</a>]. Therefore, some additional steps and costs are needed to spend on preparing samples and analyzing the global histone modifications.</p>\r\n\r\n<p>In this study, we have established a simple method (<a href=\"#figure1\">Fig. 1</a>) to isolate nuclear proteins from lesser amounts of frozen tissues. This is a very simple method that produced not only the highest percentage of histones but also carried significantly high-level housekeeping proteins.</p>\r\n\r\n<p>Concentrated protein isolated by our method and control method (acid extraction), was measured and compared by Bradford reagent as presented in <a href=\"#Table-1\">Table 1</a>. From the same amount of tissues (150mg/sample) and equal volume of dilution buffer (200ul/sample), our method yielded higher protein concentration than the acid extraction around 13 times or 29 times for inguinal fat (ING) or liver, respectively. But, the protein concentration was almost the same in the soluble (So) and nuclear (Nu) part of a homogenized sample isolated using our method. In our method, total proteins were collected with enriched histones in the nuclear part, whereas acid extraction tends to get pure nuclear proteins. However, in order to test the global histone modifications by western blot, the acid extraction method shows some weaknesses such as 1) it requires large amount of tissues for getting enough histones, 2) cannot run western blots with internal controls, and 3) isolated histones cannot be stored for a long period of time because they were extracted by 0.2N HCl. With such high protein concentration, our method showing low expressed acetylated histone 3. This result indicated that chromatins were abundant in the nucleus allowed us to run several western blots to check global modifications for all interested histones.</p>\r\n\r\n<div id=\"figure1\">\r\n<figure class=\"image\"><img alt=\"\" height=\"432\" src=\"/media/article_images/2023/14/23/178-1513778843-Figure1.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 1. </strong>Our method to isolate soluble and nuclear proteins. Soluble and nuclear proteins were isolated using ice-cold radioimmune precipitation assay buffer (RIPA) modified from a protocol we had done [<a href=\"#r-6\">6</a>-<a href=\"#r-8\">8</a>]. RIPA buffer includes 50 mM Tris, 150 mM NaCl, 1 mM EDTA, 0.1% SDS, 1% Nonodet P-40 (NP40), 0.25% sodium deoxycholate, and 0.02% sodium azide in ddH2O. 150mg of frozen mouse inguinal fat tissues (ING) or frozen liver was homogenized in 1ml of ice-cold RIPA supplemented with phenylmethylsulfonyl fluoride (PMSF – Sigma, P7626), protease inhibitor cocktail (Sigma, P8340) and PhosStop (PhosStop – Roche, 4906837001) (1). Homogenized samples were incubated on ice for 1h (2), and sonicated by VibraCell Sonicator at 40% of the sonicator’s amplitude with 3 times for 1 min (3), and then centrifuged at 10000g for 15 min at 4ºC (4). Upper liquid phase (Soluble – So) was transferred into a new 1.5ml Eppendorf for membrane and plasma proteins (5a). Pellet plus around 200µl under liquid phase (Nuclear – Nu) was kept for nuclear proteins (5b). In case of fat tissues, the fat fraction on the top of samples was carefully removed after centrifuging. Pellet was dissolved in under liquid phase by pipetting. In the control method, histone extraction was done from 150mg of frozen ING and liver of mice according to the Abcam’s protocol (http://www.abcam.com/protocols/histone-extraction-protocol-for-western-blot) using Triton Extraction Buffer, this buffer contains PBS 1X, 0.5% Triton X100 (V/V), 2mM PMSF, 0.02% Sodium azide (W/v) (NaN3). Histones were extracted by 0.2N HCl over night at 4°C (Acid). Protein concentration in both methods was measured using Bradford reagent (Sigma, B6916). </figcaption>\r\n</figure>\r\n</div>\r\n\r\n<p>In order to confirm the enrichment of nuclear proteins are isolated by our method, we performed the western blot using a specific antibody against acetylated histone 3, has been considered as one of the most common chromatins. As shown in the <a href=\"#figure2\">Fig.2</a>, the nuclear parts (Nu) of both inguinal fat (ING) (lane 2) and liver (lane 4) showing strong acetylated histone 3 expression, whereas the soluble parts (So) (lane 1 or 3) from same samples showing low expressed acetylated histone 3. This result indicated that chromatins were abundant in the nucleus allowed us to run several western blots to check global modifications for all interested histones.</p>\r\n\r\n<p>Parts (Nu) isolated by our method, even we had similar protein concentrations in the Nu and So parts of the samples (Table 1). The expression of acetylatedhistone 3 was also found from the pure histones (lane 5, 6 and 7) purified by acid extraction method, because of low protein concentration was given by this method. We were unable to load more than 20µg protein/gel-well to get stronger signals, this is one of the weaknesses of acid extraction method.</p>\r\n\r\n<div id=\"Table-1\">\r\n<p><strong><a href=\"https://jabet.bsmiab.org/table/178-1513778843-table1/\">Table-1</a>Table 1. </strong>Protein concentration of different samples.</p>\r\n</div>\r\n\r\n<p>In the immunoblotting assay, it is better to have internal controls to confirm the equal amount of loaded proteins to normalize the quantified data. Internal controls (normalization controls) are the proteins of housekeeping genes such as β actin, glyceraldehyde-3-phosphate dehydrogenase (GAPDH), hypozanthine phosphoribosyltransferase 1 (HPRT1), and ribosomal protein large P1 (RPLP1) [<a href=\"#r-11\">11</a>, <a href=\"#r-12\">12</a>]. Westernblots are used to study histones or their modifications use coomassie blue-stained gels to demonstrate equal loading [<a href=\"#r-3\">3</a>, <a href=\"#r-5\">5</a>]. In our protein isolation method, we collected the part of samples having high chromatins, so our samples still had nuclear proteins as well as proteins of housekeeping genes. Therefore, this method allowed us to have the internal controls, such as β-actin in the blot (Fig. 2). As shown in Fig. 2, from lane (1) to (4); the signals of β-actin were strong and equal in both So and Nu parts of one tissue, and in both fat (ING) and liver samples. As expected, acid extraction methods did not give any signal of β-actin in both fat and liver samples (lane 5 to 7 of Fig. 2).</p>\r\n\r\n<div id=\"figure2\">\r\n<figure class=\"image\"><img alt=\"\" height=\"272\" src=\"/media/article_images/2023/14/23/178-1513778843-Figure2.jpg\" width=\"527\" />\r\n<figcaption><strong>Figure 2. Our protein isolation method gave strong signals of acetylated histone 3 and internal control (β-actin).</strong> Acetylated histone 3 (H3) and β-actin (B. Actin) in protein isolated by our and control methods were detected by western blot as described [<a href=\"#r-9\">9</a>, <a href=\"#r-10\">10</a>, <a href=\"#r-6\">6</a>]. Blots were incubated with antibodies against acetyl histone 3 (rabbit anti acetyl H3 – Millipore, Cat. # 06-599) (1:1000), β actin (mouse anti β actin –Abcam, Ab6276) (1:10000). Specific antibody-antigen complexes were detected using fluorescent-labeled secondary antibodies (goat anti-rabbit IRDye 800, Rockland, 611-132-122; goat anti-mouse IRDye 800, Rockland, 610-730-124). Bands were visualized and quantified using the Odyssey imaging system (Licor Bioscience). </figcaption>\r\n</figure>\r\n</div>\r\n\r\n<p>The results of the current study indicate that this alternative protein isolation method is a better choice to study chromatins and other proteins as well. Moreover, this method offers several advantages including small amount of tissue samples giving high protein concentration, one gel for both targeted and control proteins, and it was simple. However, this method is only suitable for studying global expressions of proteins by western blot not by other methods such as ChiP assay as it does not yield pure chromatins.</p>"
},
{
"section_number": 2,
"section_title": "CONFLICTS OF INTEREST",
"body": "<p>The authors declare that they have no conflict of interest.</p>"
}
],
"figures": [
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/14/23/178-1513778843-Figure1.jpg",
"caption": "Figure 1. Our method to isolate soluble and nuclear proteins. Soluble and nuclear proteins were isolated using ice-cold radioimmune precipitation assay buffer (RIPA) modified from a protocol we had done [6-8]. RIPA buffer includes 50 mM Tris, 150 mM NaCl, 1 mM EDTA, 0.1% SDS, 1% Nonodet P-40 (NP40), 0.25% sodium deoxycholate, and 0.02% sodium azide in ddH2O. 150mg of frozen mouse inguinal fat tissues (ING) or frozen liver was homogenized in 1ml of ice-cold RIPA supplemented with phenylmethylsulfonyl fluoride (PMSF – Sigma, P7626), protease inhibitor cocktail (Sigma, P8340) and PhosStop (PhosStop - Roche, 4906837001) (1). Homogenized samples were incubated on ice for 1h (2), and sonicated by VibraCell Sonicator at 40% of the sonicator’s amplitude with 3 times for 1 min (3), and then centrifuged at 10000g for 15 min at 4ºC (4). Upper liquid phase (Soluble - So) was transferred into a new 1.5ml Eppendorf for membrane and plasma proteins (5a). Pellet plus around 200µl under liquid phase (Nuclear - Nu) was kept for nuclear proteins (5b). In case of fat tissues, the fat fraction on the top of samples was carefully removed after centrifuging. Pellet was dissolved in under liquid phase by pipetting. In the control method, histone extraction was done from 150mg of frozen ING and liver of mice according to the Abcam’s protocol (http://www.abcam.com/protocols/histone-extraction-protocol-for-western-blot) using Triton Extraction Buffer, this buffer contains PBS 1X, 0.5% Triton X100 (V/V), 2mM PMSF, 0.02% Sodium azide (W/v) (NaN3). Histones were extracted by 0.2N HCl over night at 4°C (Acid). Protein concentration in both methods was measured using Bradford reagent (Sigma, B6916).",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/14/23/178-1513778843-Figure2.jpg",
"caption": "Figure 2. Our protein isolation method gave strong signals of acetylated histone 3 and internal control (β-actin). Acetylated histone 3 (H3) and β-actin (B. Actin) in protein isolated by our and control methods were detected by western blot as described [9, 10, 6]. Blots were incubated with antibodies against acetyl histone 3 (rabbit anti acetyl H3 – Millipore, Cat. # 06-599) (1:1000), β actin (mouse anti β actin –Abcam, Ab6276) (1:10000). Specific antibody-antigen complexes were detected using fluorescent-labeled secondary antibodies (goat anti-rabbit IRDye 800, Rockland, 611-132-122; goat anti-mouse IRDye 800, Rockland, 610-730-124). Bands were visualized and quantified using the Odyssey imaging system (Licor Bioscience).",
"featured": true
}
],
"authors": [
{
"id": 23,
"affiliation": [
{
"affiliation": "Faculty of Biology, Hanoi National University of Education, Hanoi, Vietnam"
}
],
"first_name": "Dinh-Toi",
"family_name": "Chu",
"email": "chudinhtoi.hnue@gmail.com",
"author_order": 1,
"ORCID": "https://orcid.org/0000-0002-4596-2022",
"corresponding": true,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "Dinh-Toi Chu, \r\nFaculty of Biology, Hanoi National University of Education, Viet Nam\r\ne-mail: chudinhtoi.hnue@gmail.com",
"article": 19
},
{
"id": 24,
"affiliation": [
{
"affiliation": "College of Food Science and Technology, Nanjing Agricultural University, Nanjing 8 210095, China"
}
],
"first_name": "Yang",
"family_name": "Tao",
"email": null,
"author_order": 2,
"ORCID": "https://orcid.org/0000-0001-5276-8546",
"corresponding": false,
"co_first_author": false,
"co_author": true,
"corresponding_author_info": "",
"article": 19
}
],
"views": 1545,
"downloads": 180,
"references": [
{
"id": 31,
"serial_number": 1,
"pmc": null,
"reference": "Patterson K, Molloy L, Qu W, Clark S. DNA Methylation: Bisulphite Modification and Analysis. J Vis Exp. 2011:e3170.",
"DOI": null,
"article": 19
},
{
"id": 32,
"serial_number": 2,
"pmc": null,
"reference": "Bannister AJ, Kouzarides T. Regulation of chromatin by histone modifications. Cell Res. 2011;21:381-95.",
"DOI": null,
"article": 19
},
{
"id": 33,
"serial_number": 3,
"pmc": null,
"reference": "Trojer P, Zhang J, Yonezawa M, Schmidt A, Zheng H, Jenuwein T, et al. Dynamic Histone H1 Isotype 4 Methylation and Demethylation by Histone Lysine Methyltransferase G9a/KMT1C and the Jumonji Domain-containing JMJD2/KDM4 Proteins. Journal of Biological Chemistry. 2009;284:8395-405.",
"DOI": null,
"article": 19
},
{
"id": 34,
"serial_number": 4,
"pmc": null,
"reference": "Wellen KE, Hatzivassiliou G, Sachdeva UM, Bui TV, Cross JR, Thompson CB. ATP-Citrate Lyase Links Cellular Metabolism to Histone Acetylation. Science. 2009;324:1076-80.",
"DOI": null,
"article": 19
},
{
"id": 35,
"serial_number": 5,
"pmc": null,
"reference": "Zhang Q, Ramlee MK, Brunmeir R, Villanueva CJ, Halperin D, Xu F. Dynamic and distinct histone modifications modulate the expression of key adipogenesis regulatory genes. Cell Cycle. 2012;11:4310-22.",
"DOI": null,
"article": 19
},
{
"id": 36,
"serial_number": 6,
"pmc": null,
"reference": "Chu DT, Malinowska E, Gawronska-Kozak B, Kozak LP. Expression of Adipocyte Biomarkers in a Primary Cell Culture Models Reflects Preweaning Adipobiology. Journal of Biological Chemistry, 2014; 289(26): 18478-88.",
"DOI": null,
"article": 19
},
{
"id": 37,
"serial_number": 7,
"pmc": null,
"reference": "Jura M, Jarosławska J, Chu DT, Kozak LP. Mest and Sfrp5 are biomarkers for healthy adipose tissue. Biochimie, 2016; 124: 124-33.\r\n[8] Chu DT, Malinowska E, Jura M, Kozak LP. C57BL/6J mice as a polygenic developmental model of diet-induced obesity. Physiological Reports, 2017;5(7): 20.",
"DOI": null,
"article": 19
},
{
"id": 42,
"serial_number": 8,
"pmc": null,
"reference": "Chu DT, Malinowska E, Jura M, Kozak LP. C57BL/6J mice as a polygenic developmental model of diet-induced obesity. Physiological Reports, 2017;5(7): 20.",
"DOI": null,
"article": 19
},
{
"id": 38,
"serial_number": 9,
"pmc": null,
"reference": "Rim JS, Mynatt RL, and Gawronska-Kozak B. Mesenchymal stem cells from the outer ear: a novel adult stem cell model system for the study of adipogenesis. The FASEB Journal, 2005.",
"DOI": null,
"article": 19
},
{
"id": 39,
"serial_number": 10,
"pmc": null,
"reference": "Anunciado-Koza R, Ukropec J, Koza RA, Kozak LP. Inactivation of UCP1 and the glycerol phosphate cycle synergistically increases energy expenditure to resist diet-induced obesity. J Biol Chem, 2008. 283(41): 27688-97.",
"DOI": null,
"article": 19
},
{
"id": 40,
"serial_number": 11,
"pmc": null,
"reference": "Aldridge GM, Podrebarac DM, Greenough WT, Weiler IJ. The use of total protein stains as loading controls: An alternative to high-abundance single-protein controls in semi-quantitative immunoblotting. Journal of Neuroscience Methods. 2008;172:250-4.",
"DOI": null,
"article": 19
},
{
"id": 41,
"serial_number": 12,
"pmc": null,
"reference": "Gilda JE, Gomes AV. Stain-Free total protein staining is a superior loading control to β-actin for Western blots. Analytical Biochemistry. 2013;440:186-8.",
"DOI": null,
"article": 19
}
]
},
{
"id": 16,
"slug": "178-1513778846-dna-fingerprinting-and-molecular-diversity-analysis-for-the-improvement-of-brinjal-solanum-melongena-l-cultivars",
"featured": false,
"slider": false,
"issue": "Vol1 Issue1",
"type": "original_article",
"manuscript_id": "178-1513778846",
"recieved": "2017-11-25",
"revised": null,
"accepted": "2017-12-15",
"published": "2018-01-05",
"pdf_file": "https://jabet.bsmiab.org/media/pdf_file/2023/11/178-1513778846.pdf",
"title": "DNA fingerprinting and molecular diversity analysis for the improvement of brinjal (Solanum melongena L.) cultivars",
"abstract": "<p>An investigation was conducted to assess genetic diversity of Bangladesh Agricultural Research Institute (BARI) released 11 varieties of brinjal (BARI begun 1, BARI begun 4-10, BARI hybrid begun 1, 3 and 4) using PCR-based randomly amplified polymorphic DNA (RAPD) markers. A total of 44 distinct DNA amplified bands were observed for five primers (OPB-04, OPB-08, OPD-02, OPP-13 and OPW-08) with an average of 5.6 polymorphic bands per primer. The overall gene diversity was detected 0.216 and level of polymorphism was found 63.64%. The pair wise inter-variety similarity indices ranged from 67.03% to 97.61%. The UPGMA dendrogram segregated 11 genotypes of brinjal into two main clusters. The first major cluster had only one genotype (BARI begun 6) and the second major cluster had rest of ten genotypes. BARI begun 6 vs BARI begun 1 showed the highest Nei’s genetic distance (0.5261) as they are released from different parental origin. On the other hand, BARI begun 9 vs BARI begun 7 varietal pair showed lowest genetic distance (0.0465) as they are released from same parental origin. The experiment reveals genetic diversity and relatedness among 11 brinjal genotypes which may be informative for the future varietal identification and genetic improvement of this vegetable crop.</p>",
"journal_reference": "J Adv Biotechnol Exp Ther. 2018; 1(1) : 01-06.",
"academic_editor": "Md Saidur Rahman, PhD\r\nChung-Ang University, South Korea.",
"cite_info": "Sultana S, Islam MN, Hoque ME. DNA fingerprinting and molecular diversity analysis for the improvement of brinjal (Solanum melongena L.) cultivars. J Adv Biotechnol Exp Ther. 2018; 1(1) : 01-06.",
"keywords": [
"Genetic diversity",
"DNA fingerprinting",
"Solanum melongena"
],
"DOI": "10.5455/jabet.d1",
"sections": [
{
"section_number": 1,
"section_title": "INTRODUCTION",
"body": "<p>Brinjal (<em>Solanum melongena </em>L.) is an economically important Solanaceous vegetable, widely consumed in Asia, Europe, Africa and America [<a href=\"#r-1\">1</a>; <a href=\"#r-2\">2</a>; <a href=\"#r-3\">3</a>]. It has the early European name ‘eggplant” locally known as “Begun” in Bangladesh. It is a self-pollinated, diploid (2n=24) annual crop belongs to the family <em>Solanaceae</em>. Brinjal is largely cultivated in almost all districts of Bangladesh. The total area of eggplant cultivation is 76370 acres with total annual production of 310354 metric tons with an average yield of 4064 kg per acre in 2014-2015 [<a href=\"#r-4\">4</a>]. Globally, the eggplant is ranked as third most important crop from <em>Solanaceae</em> family after potato and tomato with an annual production of more than 41×10<sup>6</sup> tons [<a href=\"#r-5\">5</a>].<br />\r\nBrinjal is nutritious vegetable and has got multifarious use as a dish item. It is a valuable source of minerals (particularly iron) and vitamins. The fruit and other parts of the plant are used in traditional medicine [<a href=\"http://3r-6\">6</a>]. Eggplant is ranked as one of the top ten vegetables due to its high oxygen radical scavenging capacity [<a href=\"#r-7\">7</a>]. Research studies have shown that phenolic compounds of eggplant have antioxidant properties preventing oxidation and diabetes complications [<a href=\"#r-8\">8</a>; <a href=\"#r-9\">9</a>]. Additionally, the eggplant peel is rich in anthocyanins having therapeutic potential against hyperlipidemia and cardiovascular diseases by inhibiting lipid peroxidation [<a href=\"#r-9\">9</a>; <a href=\"#r-10\">10</a>].<br />\r\nGenetic resources management of brinjal cultivars is co-related with the sustainable agricultural and economic development of the country. The yield potential of this eggplant in Bangladesh is very low compared to other countries due to the incidence of insect pests and diseases [<a href=\"#r-11\">11</a>; <a href=\"#r-12\">12</a>]. Molecular characterization of this eggplant may provide an avenue to screen the germplasm resources for any selective breeding and genetic improvement. Molecular markers are reliable tools to characterize the DNA profile of plant genotypes to study the genetic diversity. In Bangladesh, genetic data on brinjal is not rich enough. So genetic status of this important crop is needed to be established and documented by using DNA markers which may provide valuable information for further breeding programme. Randomly amplified polymorphic DNA (RAPD) markers are effective for the evaluation of genetic diversity due to easy application and relatively low expensive compared to other molecular markers [<a href=\"#r-13\">13</a>; <a href=\"#r-14\">14</a>]. Molecular characterization by RAPD markers is easy and rapid. RAPD markers are being used for the identification of genetic relationship among cultivars [<a href=\"#r-15\">15</a>; <a href=\"#r-16\">16</a>].<br />\r\nThe present investigation was formulated with the objective of studying genetic variation among the different released brinjal cultivars through RAPD markers, molecular characterization by DNA profiling and <strong>t</strong>o reveal phylogenetic relatedness and genetic distance among the brinjal genotypes.</p>"
},
{
"section_number": 2,
"section_title": "MATERIALS AND METHODS",
"body": "<p><strong>Experimental plant materials and their sources</strong><br />\r\nThe plant materials for this study consisted of 11 brinjal varieties namely BARI begun 1, BARI begun 4-10, F1 hybrids (BARI hybrid begun 1, 3 and 4). All the varieties were collected from Horticulture Research Centre (HRC) of Bangladesh Agricultural Research Institute (BARI), Joydebpur, Gazipur. Seedlings were grown at the Laboratory of Department of Biotechnology at Sher-e-Bangla Agricultural University, Dhaka in earthen tubs containing soils collected from nursery.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Genomic DNA isolation</strong><br />\r\nGenomic DNA of the brinjal seedlings was isolated following protocol described by Saghai-Maroof and co-workers [<a href=\"#r-17\">17</a>] as well as Islam and co-workers [<a href=\"#r-18\">18</a>; <a href=\"#r-19\">19</a>] with some modifications. Approximately 200 mg of young leaves of 30 days old plants were cut into minuscule pieces, homogenized and digested with extraction buffer (pH= 8.0): 50 mM Tris-HCl, 25 mM EDTA (Ethylene­diaminetetraacetic acid), 300 mM NaCl and 1% SDS (Sodium Dodecyl Sulfate). Following incubation for 20 minutes at 65°C with intermittent shaking, the mixture was vortexed with an equal volume of phenol: chloroform: isoamyl alcohol (25:24:1, v/v/v) for 20 seconds. The emulsified mixture was centrifuged at 13000 rpm for 10 minutes. The supernatant was used for precipitation with 2/3 volume of 100% alcohol in presence of 0.3 M sodium acetate and DNA was pelleted by centrifugation. The DNA pellets were then washed with 70% ethanol. The air dried DNA pellets were resuspended in an appropriate volume of TE buffer (10 mM Tris-HCl, 1 mM EDTA, pH=8.0) and finally treated with RNAse and stored at -20°C. The quality and quantity of DNA were checked by electrophoresis and spectrophotometer, respectively.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Screening and selection of RAPD primers</strong><br />\r\nInitially, nine decamer primers namely OPA-18, OPB-04, OPB-06, OPB-08, OPD-02, OPF-08, OPG-19, OPP-13 and OPW-08 (Operon Technologies, Inc. Alameda, California, USA) were selected and screened by polymerase chain reaction (PCR) for their ability to produce polymorphic patterns in brinjal varieties and five primers (OPB-04, OPB-08, OPD-02, OPP-13 and OPW-08) which gave reproducible and distinct polymorphic amplified products were selected finally for RAPD analysis.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Polymerase chain reaction</strong><br />\r\nPCR reactions were performed for each DNA sample in a 25 µL reaction mix containing 12.5 µL 2X Taq Master mix (GeneON, Germany), 2.5 µL of RAPD primer (10 pM), 7.5 µL of sterile de-ionized water and 2.5 µL of genomic DNA (25ng/ µL). DNA amplification was performed in an oil-free thermal cycler by setting a thermal profile as pre-heating at 95o C for 5 min followed by 33 cycles of 45 s denaturation at 95o C, 30 s annealing at 30o C and extension at 72o C for 1 min. After the last cycle, a final step of 5 min at 72o C was added to allow complete extension of all amplified fragments.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Visualization of PCR products</strong><br />\r\nPCR products were confirmed by electrophoresis on 1.5 % agarose gel containing 1 µL ethidium bromide (10 mg/L) in 1X TBE buffer at 85 V for 50 min. After electrophoresis, the gel was taken out carefully from the gel chamber and was placed on high-performance ultraviolet light box (UV trans-illuminator) of gel documentation for checking the RAPD bands and photographed by a Gel Cam Polaroid camera.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>RAPD data analysis</strong><br />\r\nFollowing electrophoresis, the sizes of the PCR products were estimated by comparisons of distance travelled by each fragment with distance travelled by known size fragments of the DNA molecular weight markers (100 bp DNA ladder, Genei, India or 1 Kb DNA ladder, BRIC, Korea). All distinct bands or fragments (RAPD markers) were thereby given identification numbers according to size and scored visually on the basis of their presence (1) or absence (0), separately for each variety for each primer. The scores recorded for all RAPD primers were then pooled to create a single data matrix. This was used to compare the frequencies of all polymorphic RAPD markers among populations with 1000 simulated samples using POPGENE (version 1.31) [<a href=\"#r-20\">20</a>] computer program. The size of the RAPD markers were estimated by using the software DNAfrag, version 3.03 [<a href=\"#r-21\">21</a>]. Nei’s [<a href=\"#r-22\">22</a>] genetic distance values were calculated based on the frequencies of polymorphic markers using the POPGENE (Version 1.31) computer package [<a href=\"#r-20\">20</a>]. For each population group, Nei’s gene diversity (h) was calculated for each locus and then averaged over all loci.<br />\r\nThese parameters were performed using POPGENE software [<a href=\"#r-20\">20</a>] in the formula: h=1-∑pi2, I= ∑pi log pi, where pi = the frequency of the allele i in the population. A value indicates small genetic distances, 0.10-0.15 indicates moderate genetic distances, 0.15-0.2 indicates high genetic distances and >0.2 indicates very large genetic distances [<a href=\"#r-23\">23</a>]. The dendrogram was constructed using POPGENE (Version 1.31) [<a href=\"#r-20\">20</a>]. Genetic similarity values were calculated manually based on the formula, Similarity Index (SI) = 2N­­­­­XY/(NX + NY); where, NXY is the number of RAPD bands shared by individuals X and Y, respectively, and NX and NY are the number of bands in individuals X and Y, respectively [<a href=\"#r-24\">24</a>; <a href=\"#r-25\">25</a>].</p>"
},
{
"section_number": 3,
"section_title": "RESULTS",
"body": "<p style=\"text-align:justify\"><strong>Profiling of RAPD bands and their polymorphisms</strong><br />\r\nA total of 44 distinct and different PCR based RAPD bands were produced with 8.8 per primer. The size of the bands ranges from 198 to 2898 bp (<a href=\"#Table-1\">Table 1</a>). The highest number of bands (12) was generated by primer OPD-02. Besides, the primer OPB-04, OPB-08, OPP-13 and OPW-08 generated 8 scorable bands each. The RAPD banding profile is shown in <a href=\"#figure1\">Figure 1</a>. The highest proportion of polymorphic bands (87.5%) was detected by primer OPW-08 while primer OPB-04 detected the least proportion (37.5%) and overall level of polymorphism was 63.64% (<a href=\"#Table-1\">Table 1</a>). The average values of Nei’s gene diversity for the RAPD primers ranged from 0.083 to 0.388 with an overall value of 0.216 (<a href=\"#Table-1\">Table 1</a>).</p>\r\n\r\n<div id=\"Table-1\">\r\n<p><a href=\"https://jabet.bsmiab.org/table/178-1513778846-1/\">Table-1</a><strong>Table 1.</strong> RAPD primers with corresponding number and size range of bands together with a percentage of polymorphic loci and Nei‟s gene diversity values observed in 11 brinjal varieties.</p>\r\n</div>\r\n\r\n<div id=\"figure1\">\r\n<figure class=\"image\"><img alt=\"\" height=\"752\" src=\"/media/article_images/2024/14/08/Figure1.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 1</strong>. DNA fingerprinting profile of 11 brinjal varieties based on five RAPD primers, (A) OPB-04, (B) OPB-08, (C) OPD-02, (D) OPP-13 and (E) OPW-08. Lane 1: BARI begun 1, lane 2: BARI begun 4, lane 3: BARI begun 5, lane 4: BARI begun 6, lane 5: BARI begun 7, lane 6: BARI begun 8, lane 7: BARI begun 9, <strong>l</strong>ane 8: BARI begun 10, lane 9: BARI hybrid begun 1, lane 10: BARI hybrid begun 3, lane 11: BARI hybrid begun 4. Ms: M1=100 bp DNA ladder and M2: 1 kb DNA ladder.</figcaption>\r\n</figure>\r\n</div>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Inter-variety similarity indices</strong><br />\r\nInter-varietal similarity indices (Sij) ranged from 67.03% to 97.61% (Data are not shown due to large sized table). The highest similarity indices (97.61%) were found between BARI begun 7 and BARI begun 9. BARI begun 6 and BARI begun 10 showed least inter-variety similarity indices 67.03%. All the 55 varietal pairs were not homogenous at different number of loci.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Genetic identity and genetic distance</strong><br />\r\nThe highest Nei’s genetic identity (0.9545) was observed in BARI begun 9 vs BARI begun 7 varietal pairs whereas the lowest genetic identity (0.6136) was estimated in BARI begun 6 vs BARI begun 5 varietal pair (<a href=\"#Table-2\">Table 2</a>). The result indicates that the low or high level genetic distance exists between varieties with their same or different origins. BARI began 6 vs BARI begun 1 showed highest Nei’s genetic distance (0.5261) as they are released from different parental origin. On the other hand BARI began 9 vs BARI begun 8 varietal pair showed lowest genetic distance (0.0706) as they are released from same parental origin. This variation might be created due to geographical origin.</p>\r\n\r\n<div id=\"Table-1\">\r\n<p><a href=\"https://jabet.bsmiab.org/table/178-1513778846-table2/\">Table-2</a><strong>Table 2.</strong> Neil's genetic identity.</p>\r\n</div>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Phylogenetic relatedness and construction of dendrogram</strong><br />\r\nPhylogenetic dendrogram based on Nei’s (1972) genetic distance using Unweighted Pair Group Method of Arithmetic Means (UPGMA) indicated the segregation of 11 genotypes of brinjal into two main clusters, ‘A’ and ‘B’ (<a href=\"#figure2\">Figure 2</a>). The first major cluster ‘A’ had only one genotype (BARI begun 6) and the second major cluster ‘B’ had rest of ten genotypes. The second major cluster subdivided into two minor clusters (C & D) in which one cluster ‘D’ had only one genotype (BARI begun 1) and the other cluster ‘C’ had rest of the nine varieties. This minor cluster further subdivided into two clusters (E & F). In which cluster E divided into two sub clusters (G and H). BARI hybrid begun 1 formed cluster ‘G’ and BARI begun 7, BARI begun 8 and BARI begun 9 grouped in cluster ‘H’. Cluster ‘I’ and ‘J’ were the subdivision of cluster ‘F’. BARI hybrid begun 3 and BARI hybrid begun 4 formed cluster ‘I’ and BARI begun 4, BARI begun 5 and BARI begun 10 were grouped in cluster ‘J’.</p>\r\n\r\n<div id=\"figure2\">\r\n<figure class=\"image\"><img alt=\"\" height=\"341\" src=\"/media/article_images/2024/14/08/Figure2.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 2.</strong> The UPGMA (Unweighted pair group method of arithmetic mean) dendrogram based on Nei‟s (1972) genetic distance within 11 brinjal varieties according to RAPD analysis.</figcaption>\r\n</figure>\r\n</div>"
},
{
"section_number": 4,
"section_title": "DISCUSSION",
"body": "<p>With advancement of DNA marker technology, molecular characterization plays a vital role in managing and utilizing plant genetic resources [<a href=\"#r-26\">26</a>]. It is immensely helpful in selective breeding from diverse parents to widen the breeding gene pool [<a href=\"#r-27\">27</a>]. Thereby, DNA markers are increasingly adopted as an effective and appropriate tool for basic and applied studies addressing biological components in agricultural production systems [<a href=\"#r-28\">28</a>].</p>\r\n\r\n<p>In the present study, the overall level of polymorphism (63.64%) indicated the effectiveness of RAPD technique to investigate genetic diversity among the different genotypes of brinjal. Number of polymorphic bands ranged from 3-9 with an average of 5.6. Verma and co-workers [<a href=\"#r-29\">29</a>] reported 5.58 bands per primer and Islam and co-workers [<a href=\"#r-30\">30</a>] found 5.67 bands per primer in brinjal varieties which are mostly consistent with the present study. Almost similar level of polymorphisms was reported in different experiment with brinjal and other eggplants, for instance 57.89% [<a href=\"#r-31\">31</a>], 57.58% [<a href=\"#r-32\">32</a>]. In contrast, higher level of polymorphism in brinjal germplasm was obtained by Aguoru and co-workers [<a href=\"#r-33\">33</a>] (83.33%) as well as by Islam and coworkers [<a href=\"#r-30\">30</a>] (70.59%). The value of overall gene diversity was 0.216 in this study which is very close to the findings reported by Biswas and co-workers [<a href=\"#r-31\">31</a>], where they observed overall gene diversity value of 0.23 among 10 promising brinjal varieties with RAPD markers.</p>\r\n\r\n<p>Karihaloo and co-workers [<a href=\"#r-34\">34</a>] reported the highest similarity (0.947) between <em>S. insanum</em> and <em>S. melongena</em> while analyzing genetic similarity of 52 accessions of Solanum. The present study indicates a high level of genetic variation within some brinjal varieties which were perceived from the data of inter-variety similarity indices and genetic distance value. Phylogenetic clustering patterns based on genetic distance values also indicated diverse relationship that occurred might be due to geographical origin of the parental varieties.<br />\r\nFinally it can be concluded that high degree of diversity of the studied brinjal cultivars may be attributable to genetic improvement programme based on the clustering patterns. It will also provide support for selection of crossing combinations from parental genotypes and for broadening the genetic basis of breeding programs. Due to having some medicinal values particularly against cancer, diabetes and cardiovascular disease, marker assisted breeding and production of this important eggplant may contribute to enrich diets and bring health benefits.</p>"
},
{
"section_number": 5,
"section_title": "ACKNOWLEDGEMENT",
"body": "<p>The authors express thanks to the Ministry of Science and Technology, Government of the Peoples’ Republic of Bangladesh for providing NST (National Science and Technology) fellowship with financial support to conduct this fundamental research.</p>"
},
{
"section_number": 6,
"section_title": "AUTHOR CONTRIBUTIONS",
"body": "<p>SS performed the experiment; MNI and MEH conceived the study; SS and MNI analyzed the data; SS and MNI wrote the paper.</p>"
},
{
"section_number": 7,
"section_title": "CONFLICT OF INTERESTS",
"body": "<p>The authors declare no conflict of interest.</p>"
}
],
"figures": [
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/14/08/Figure1.jpg",
"caption": "Figure 1. DNA fingerprinting profile of 11 brinjal varieties based on five RAPD primers, (A) OPB-04, (B) OPB-08, (C) OPD-02, (D) OPP-13 and (E) OPW-08. Lane 1: BARI begun 1, lane 2: BARI begun 4, lane 3: BARI begun 5, lane 4: BARI begun 6, lane 5: BARI begun 7, lane 6: BARI begun 8, lane 7: BARI begun 9, lane 8: BARI begun 10, lane 9: BARI hybrid begun 1, lane 10: BARI hybrid begun 3, lane 11: BARI hybrid begun 4. Ms: M1=100 bp DNA ladder and M2: 1 kb DNA ladder.",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/14/08/Figure2.jpg",
"caption": "Figure 2. The UPGMA (Unweighted pair group method of arithmetic mean)\r\ndendrogram based on Nei‟s (1972) genetic distance within 11 brinjal varieties\r\naccording to RAPD analysis.",
"featured": true
}
],
"authors": [
{
"id": 14,
"affiliation": [
{
"affiliation": "Department of Biotechnology, Sher-e-Bangla Agricultural University, Dhaka-1207, Bangladesh"
}
],
"first_name": "Sharmin",
"family_name": "Sultana",
"email": null,
"author_order": 1,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 16
},
{
"id": 15,
"affiliation": [
{
"affiliation": "Department of Biotechnology, Sher-e-Bangla Agricultural University, Dhaka-1207, Bangladesh"
}
],
"first_name": "Mohammad Nazrul",
"family_name": "Islam",
"email": "nazrul2010@sau.edu.bd",
"author_order": 2,
"ORCID": "https://orcid.org/0000-0001-5692-3566",
"corresponding": true,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "Mohammad Nazrul Islam, PhD\r\nDepartment of Biotechnology, Sher-e-Bangla Agricultural University, Dhaka-1207, Bangladesh",
"article": 16
},
{
"id": 16,
"affiliation": [
{
"affiliation": "Department of Biotechnology, Sher-e-Bangla Agricultural University, Dhaka-1207, Bangladesh"
}
],
"first_name": "Md Ekramul",
"family_name": "Hoque",
"email": null,
"author_order": 3,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": true,
"corresponding_author_info": "",
"article": 16
}
],
"views": 1191,
"downloads": 125,
"references": [
{
"id": 22,
"serial_number": 1,
"pmc": null,
"reference": "Harish DK, Agasimani AK, Imamsaheb SJ, Patil SS. Growth and yield parameters in brinjal as influenced by organic nutrient management and plant protection conditions. Res J Agril Sci 2011; 2(2): 221-25.",
"DOI": null,
"article": 16
},
{
"id": 23,
"serial_number": 2,
"pmc": null,
"reference": "Ali Z, Xu ZL, Zhang DY, He XL, Bahadur S, Yi JX. Molecular diversity analysis of eggplant (Solanum melongena) genetic resources. Genet Mol Res 2011; 10(2): 1141-55.",
"DOI": null,
"article": 16
},
{
"id": 24,
"serial_number": 3,
"pmc": null,
"reference": "Kumar G, Meena BL, Kar R, Tiwari SK. Morphological diversity in brinjal (Solanum melongena L.) germplasm accessions. Plant Genet Resour 2008; 6(3): 232-36.",
"DOI": null,
"article": 16
},
{
"id": 25,
"serial_number": 4,
"pmc": null,
"reference": "BBS. Yearbook of Agricultural Statistics-2015 (27th Series), Bangladesh Bureau of Statistics, Ministry of Planning, Government of the People’s Republic of Bangladesh, Dhaka, Bangladesh, 2016.",
"DOI": null,
"article": 16
},
{
"id": 447,
"serial_number": 5,
"pmc": null,
"reference": "Food and Agriculture Organization (FAO) of the United Nations, Agriculture database, 2010.",
"DOI": null,
"article": 16
},
{
"id": 26,
"serial_number": 6,
"pmc": null,
"reference": "Kashyap V, Kumar S, Collonier C, Fusari F, Haicour R, Rotino GL, Sihachak RD, Rajam MV. Biotechnology of eggplant. Sci Hort 2003; 97(1): 1-25.",
"DOI": null,
"article": 16
},
{
"id": 27,
"serial_number": 7,
"pmc": null,
"reference": "Jung EJ, Bae MS, Jo EK, Jo YH, Lee SC. Antioxidant activity of different parts of eggplant. J Med Plants Res 2011; 5(18): 4610-15.",
"DOI": null,
"article": 16
},
{
"id": 28,
"serial_number": 8,
"pmc": null,
"reference": "Kwon YI, Apostolidis E, Shetty K. In vitro studies of eggplant (Solanum melongena) phenolics as inhibitors of key enzymes relevant for type 2 diabetes and hypertension. Bioresour Technol 2008; 99(8): 2981-88.",
"DOI": null,
"article": 16
},
{
"id": 29,
"serial_number": 9,
"pmc": null,
"reference": "Scorsatto M, Pimentel AC, Silva AJR, Sabally K, Rosa G, Oliveira GMM. Assessment of bioactive compounds, physicochemical composition, and in vitro antioxidant activity of eggplant flour. Int J Cardiovasc Sci 2017; 30(3): 235-42.",
"DOI": null,
"article": 16
},
{
"id": 30,
"serial_number": 10,
"pmc": null,
"reference": "Basuny AM, Arafat SM, El-Marzooq MA. Antioxidant and antihyperlipidemic activities of anthocyanins from eggplant peels. J Pharma Res Rev 2012; 2(3): 50-57.",
"DOI": null,
"article": 16
},
{
"id": 43,
"serial_number": 11,
"pmc": null,
"reference": "Das AN, Singh BR. Field reaction of brinjal varieties against shoot and fruit borer, Leucinodes orbonalis. Environ Eco 2000; 8(2): 761-62.",
"DOI": null,
"article": 16
},
{
"id": 44,
"serial_number": 12,
"pmc": null,
"reference": "Rashid MM. A Guidebook of Plant Pathology. Department of Plant Pathology, Hajee Mohammad Danesh Science and Technology University, Bangladesh, 2000, p58.",
"DOI": null,
"article": 16
},
{
"id": 45,
"serial_number": 13,
"pmc": null,
"reference": "Williams JGK, Hanafey MK, Rafalski JA, Tingey SV. Genetic analysis using random amplified polymorphic DNA markers. Methods Enzymol 1990; 1218: 704-40.",
"DOI": null,
"article": 16
},
{
"id": 46,
"serial_number": 14,
"pmc": null,
"reference": "Rafalski JA, Tingey SV. Genetic diagnostics in plant breeding: RAPDs, microsatellites and machines. Trends Genet 1999; 9: 275-79.",
"DOI": null,
"article": 16
},
{
"id": 47,
"serial_number": 15,
"pmc": null,
"reference": "Afzal MA, Haque MM, Shanmugasundaram S. Random Amplified Polymorphic DNA (RAPD) analysis of selected mungbean (Vigna radiata L. Wilczek) cultivars. Asian J Plant Sci 2004; 3(1): 20-24.",
"DOI": null,
"article": 16
},
{
"id": 48,
"serial_number": 16,
"pmc": null,
"reference": "Tosti N, Negri V. Effiency of three PCR-based markers in assessing genetic variation among cowpea (vigna ungiculata spp unguiculata) landraces. Genome 2002; 45: 268-75.",
"DOI": null,
"article": 16
},
{
"id": 49,
"serial_number": 17,
"pmc": null,
"reference": "Saghai-Maroof MA, Soliman KM, Jonsensan RA, Allard RW. Ribosomal spacer length polymorphism in barley: Mendelian inheritance, chromosomal location and population dynamics. Proc Natl Acad Sci USA, 1984; 81: 8014-19.",
"DOI": null,
"article": 16
},
{
"id": 50,
"serial_number": 18,
"pmc": null,
"reference": "Islam MN, Molla MR and Rahman L. Genetic characterization of sesame varieties through RAPD fingerprinting technique in Bangladesh. J Sher-e-Bangla Agric Univ 2013; 7(1): 1-5.",
"DOI": null,
"article": 16
},
{
"id": 51,
"serial_number": 19,
"pmc": null,
"reference": "Islam MN, Molla MR, Rohman MM, Hasanuzzaman M, Islam SMN and Rahman L. DNA fingerprinting and genotyping of cotton varieties using SSR Markers. Not. Bot. Horti Agrobot. Cluj-Napoca 2012; 40: 261-65.",
"DOI": null,
"article": 16
},
{
"id": 52,
"serial_number": 20,
"pmc": null,
"reference": "Yeh FC, Yang RC, Boyle T. POPGENE VERSION 1.31: Microsoft Window-based free software for population genetic analysis. ftp://ftp.microsoft.com/Softlib/ HPGL.EXE, 1999.",
"DOI": null,
"article": 16
},
{
"id": 53,
"serial_number": 21,
"pmc": null,
"reference": "Nash JHE. DNAfrag, Version 3.03. Institute for Biological Sciences, National Research Council of Canada, Ottawa, Ontario, Canada, 1991.",
"DOI": null,
"article": 16
},
{
"id": 54,
"serial_number": 22,
"pmc": null,
"reference": "Nei M. Genetic distance between populations. Am Nat 1972; 106: 283-92.",
"DOI": null,
"article": 16
},
{
"id": 55,
"serial_number": 23,
"pmc": null,
"reference": "Nei M. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics 1978; 89: 583-90.",
"DOI": null,
"article": 16
},
{
"id": 56,
"serial_number": 24,
"pmc": null,
"reference": "Chapco W, Ashton NW, Martel RKB, Antonishyn N. A feasibility study of the use of random amplified polymorphic DNA in the population genetics and systematics of grasshoppers. Genome 1992; 35: 569-74.",
"DOI": null,
"article": 16
},
{
"id": 57,
"serial_number": 25,
"pmc": null,
"reference": "Lynch M. The similarity index and DNA fingerprinting. Mol Biol Evol 1990; 7:478-84.",
"DOI": null,
"article": 16
},
{
"id": 58,
"serial_number": 26,
"pmc": null,
"reference": "Karp A. The new genetic era: will it help us in managing genetic diversity? 43-56 p. In: Engels JMM, Rao VR, Brown AHD, Jackson MT (Eds.). Managing Plant Genetic diversity. International Plant Genetic Resources Institute, Rome, Italy, 2002.",
"DOI": null,
"article": 16
},
{
"id": 59,
"serial_number": 27,
"pmc": null,
"reference": "Fu YB. Genetics redundancy and distinctness of flax germplasm as revealed by RAPD dissimilarity. Plant Genet Resour 2006; 4:177-84.",
"DOI": null,
"article": 16
},
{
"id": 60,
"serial_number": 28,
"pmc": null,
"reference": "Jones N, Ougham H, Thomas H. Genome mapping, molecular markers and marker–assisted selection in crop plants. New phytol 1997; 137:165-77.",
"DOI": null,
"article": 16
},
{
"id": 61,
"serial_number": 29,
"pmc": null,
"reference": "Verma M, Rathi S, Munshi AD, Kumar A, Arya L, Bhat KV, Kumar R. Genetic diversity of Indian brinjal revealed by RAPD and SSR markers. Indian J Horticult 2012; 69 (4): 517-22.",
"DOI": null,
"article": 16
},
{
"id": 62,
"serial_number": 30,
"pmc": null,
"reference": "Islam Z, Siddiqua MK, Hasan MM, Nahar MA, Islam MA, Shamsuzzaman SM, Mondol MMA, Puteh AB. Assessment of genetic diversity of brinjal (Solanummelongena L.) germplasm by RAPD markers. Res Crops 2014; 15(2): 416-22.",
"DOI": null,
"article": 16
},
{
"id": 63,
"serial_number": 31,
"pmc": null,
"reference": "Biswas MS, AkhondMA,Yousuf M, Al‐Amin, Mahmuda K, Muhammed RK. Genetic relationship among ten promising eggplant varieties using RAPD markers. Plant Tissue Cult Biotech 2009; 19(2): 119-26.",
"DOI": null,
"article": 16
},
{
"id": 64,
"serial_number": 32,
"pmc": null,
"reference": "Sharmin D, Khalil MI, Begum SN, Meah MB. Molecular characterization of eggplant crosses by using RAPD analysis. Int J Sustainability Crop Prod 2011; 6: 22- 28.",
"DOI": null,
"article": 16
},
{
"id": 65,
"serial_number": 33,
"pmc": null,
"reference": "Aguoru CU, Omoigui LO, Olasan JO. Population genetic study of eggplants (Solanum) species in Nigeria, Tropical\r\nWest Africa, using molecular markers.Int J Plant Res 2015; 5: 7-12.",
"DOI": null,
"article": 16
},
{
"id": 66,
"serial_number": 34,
"pmc": null,
"reference": "Karihaloo JL, Brauner S, Gottlieb LD. Random amplified polymorphic DNA variation in the eggplant, Solanummelongena L. (Solanaceae). Theor Applied Genet. 1995; 90(6): 767-70.",
"DOI": null,
"article": 16
}
]
}
]
}
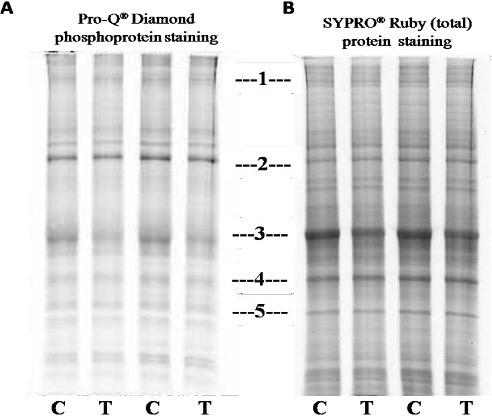{kind=link}
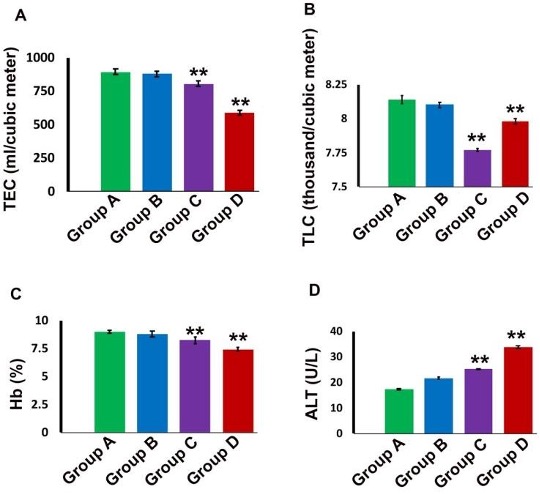{kind=link}
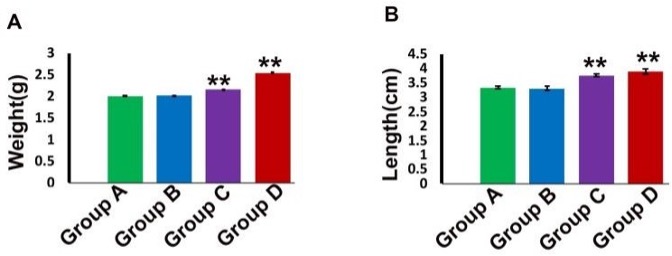{kind=link}
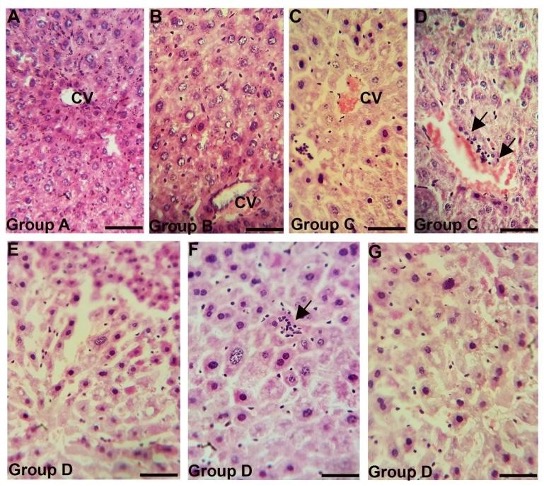{kind=link}
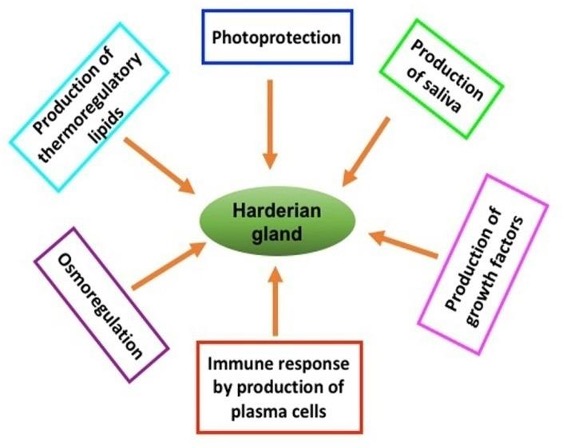{kind=link}
{kind=link}
{kind=link}
{kind=link}
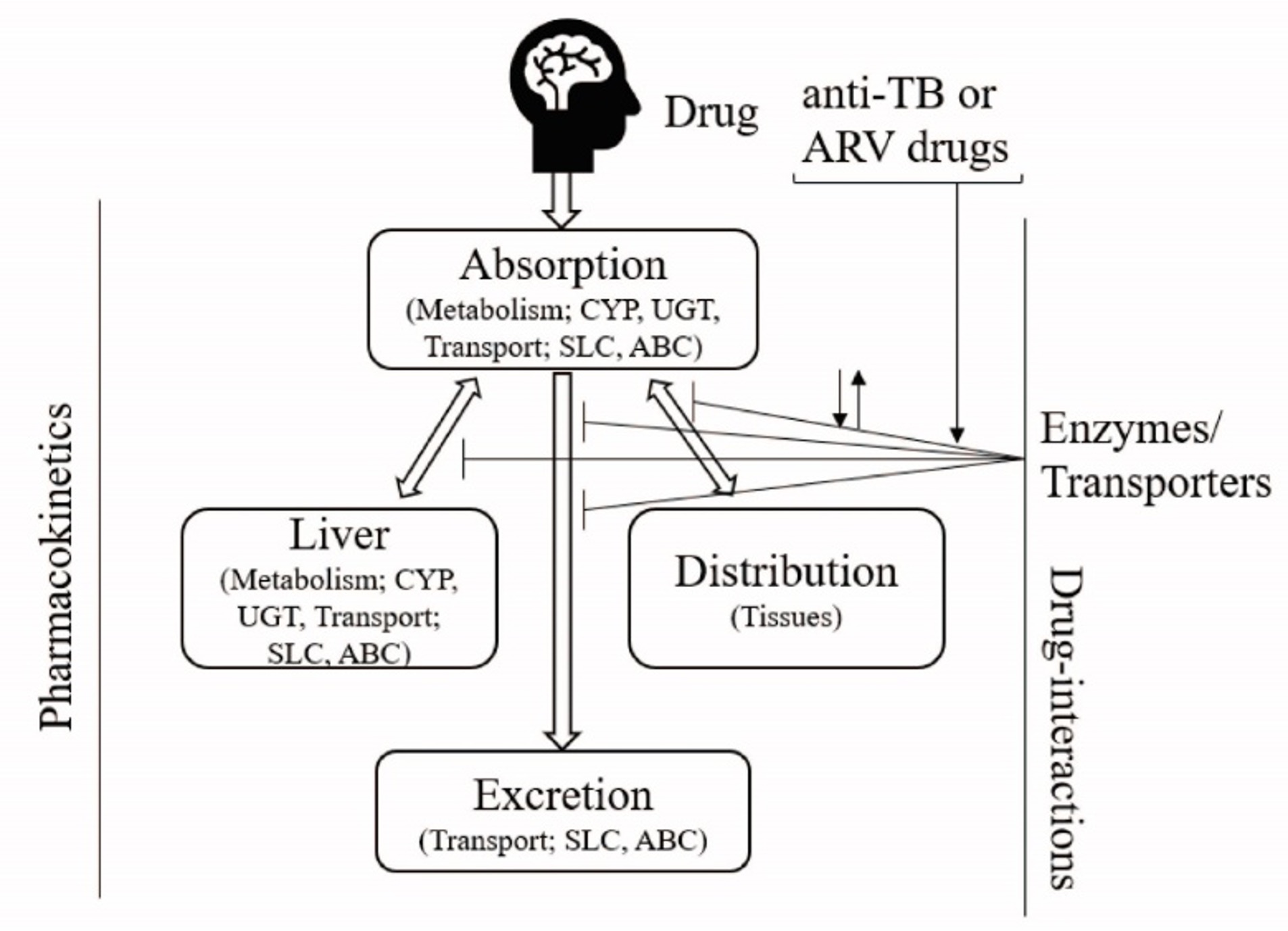{kind=link}
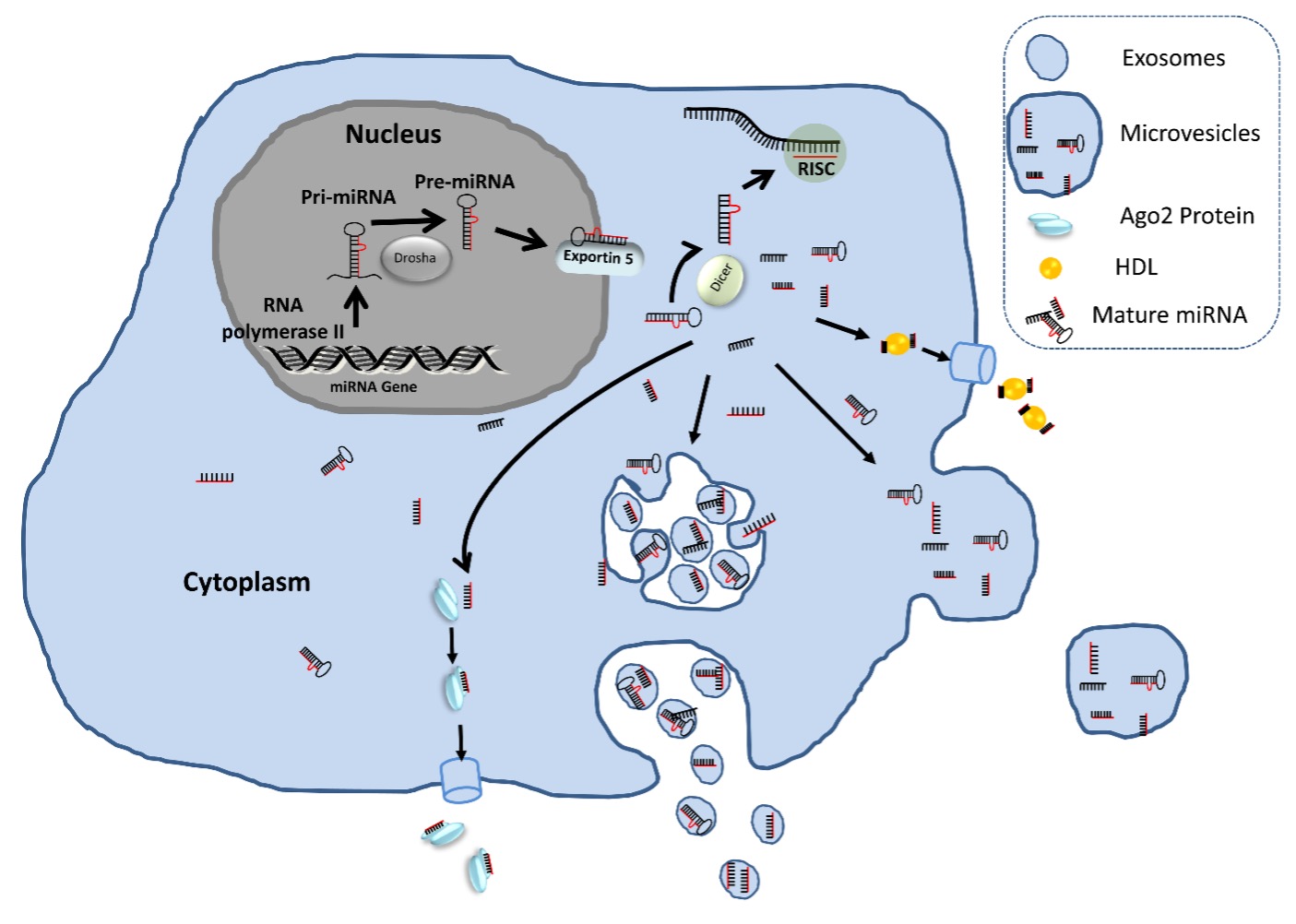{kind=link}
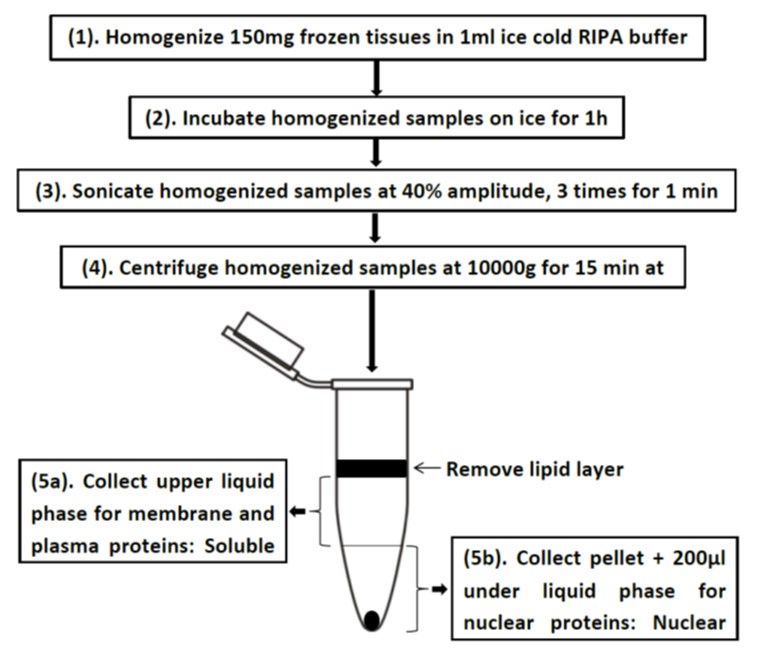{kind=link}
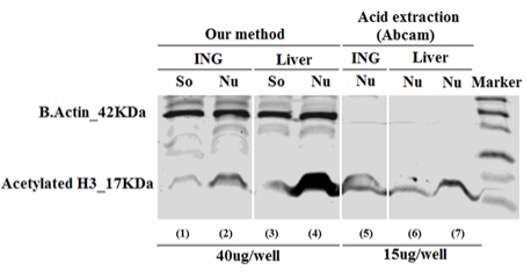{kind=link}
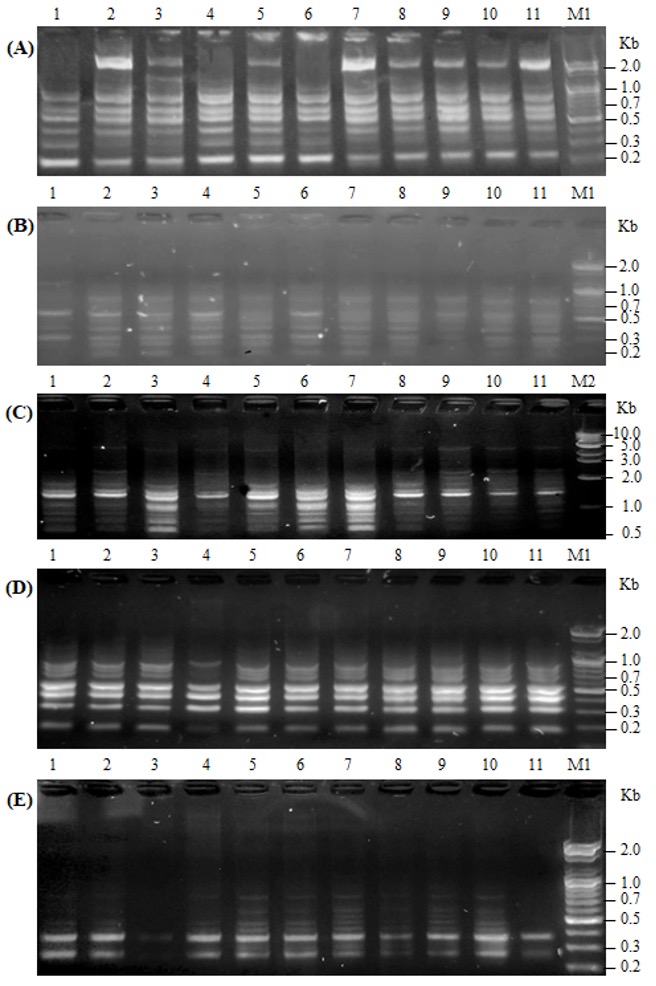{kind=link}
{kind=link}