HTTP 200 OK
Allow: GET, HEAD, OPTIONS
Content-Type: application/json
Vary: Accept
{
"count": 319,
"next": "https://jabet.bsmiab.org/articles/?format=api&page=25",
"previous": "https://jabet.bsmiab.org/articles/?format=api&page=23",
"results": [
{
"id": 130,
"slug": "178-1597659391-effect-of-chemical-and-physical-mutagens-on-amylase-producing-potentiality-of-aspergillus-flavus-nsh9",
"featured": false,
"slider": false,
"issue": "Vol4 Issue1",
"type": "original_article",
"manuscript_id": "178-1597659391",
"recieved": "2020-08-17",
"revised": null,
"accepted": "2020-10-24",
"published": "2020-10-27",
"pdf_file": "https://jabet.bsmiab.org/media/pdf_file/2023/00/178-1597659391.pdf",
"title": "Effect of chemical and physical mutagens on amylase producing potentiality of Aspergillus flavus NSH9",
"abstract": "<p><em>Aspergillus flavus</em> NSH9 is a potential source of α-amylase and glucoamylases. Between two of its glucoamylases, one can digest the raw starch granules, while both being pH and thermostable. The purpose of the study was to enhance the production of amylases (α-amylase and Glucoamylase) from <em>A. flavus</em> NSH9 by random mutagenesis using Ultraviolet (UV) irradiation and Ethidium bromide (EtBr) treatment. Glucoamylase and α-amylase activity were evaluated by 3, 5-dinitrosalicyclic acid (DNS) method and starch-iodine method, respectively. Mutated <em>A. flavus</em> NSH9 by EtBr treatment (10µg/ml) in PDA plate produced highest amount of both crude glucoamylases (GA) (1.47 ± 0.087 U/mL/min) and raw starch degrading glucoamylase (RSD-GA) (0.839 ± 0.036 U/mL/Hour). The highest value was 1.6 and 1.5 times higher compared to control for GA and RSD-GA, respectively. Here as, UV induced radiation produced about 1.3 and 1.4 times more GA and RSD-GA compared to control, respectively. The activity of α-amylase was about 1.7 times higher in the treatment group with 5µg/ml EtBr in sublethal condition than without the treatment group. In conclusion, both UV and EtBr treatment increased the amylases production from <em>A. flavus</em> NSH9. As the single mutation process of <em>A. flavus</em> NSH9 enhanced all three enzymes, the strains could be used for the commercial production of amylase.</p>",
"journal_reference": "J Adv Biotechnol Exp Ther. 2021; 4(1): 53-59.",
"academic_editor": "Md. Niamul Haque, PhD; Incheon National University, South Korea",
"cite_info": "Ruaida A, Karim KMR. Effect of chemical and physical mutagens on amylase producing potentiality of Aspergillus flavus NSH9. J Adv Biotechnol Exp Ther. 2021; 4(1): 53-59.",
"keywords": [
"Mutation",
"Glucoamylase",
"UV radiation",
"Aspergillus flavus NSH9",
"Alpha amylase"
],
"DOI": "10.5455/jabet.2021.d106",
"sections": [
{
"section_number": 1,
"section_title": "INTRODUCTION",
"body": "<p>The amylases (α-amylases, β-amylases, and glucoamylases) are among the most valuable enzymes and are important for biotechnology, representing a class of industrial enzymes with approximately 30% of the world enzyme market [<a href=\"#r-1\">1-2</a>]. Alpha amylase (1,4-α-D-glucan glucanohydrolase, E.C. 3.2.1.1) is a family of endo amylases that randomly cleaves the α-1,4 linkages throughout of the starch particles, generating glucose, maltose, dextrin, and oligosaccharides [<a href=\"#r-3\">3</a>]. Whereas, glucoamylase (1,4-α-D-glucan glucohydrolase, E.C. 3.2.1.3, GA) is an exo-acting enzyme that produces β-D-glucose from the nonreducing ends of raw or soluble starches and associated polysaccharide chains by hydrolyzing α-1, 4 linkages [<a href=\"#r-4\">4</a>]. Both amylases have a considerable role in starch processing in the food industries, for instance, for the glucose and fructose syrup derived from liquefied starch with both amylase and glucoamylase action [<a href=\"#r-5\">5-6</a>]. Amylase is also used in different food processing, beverage, various fermented foods, pharmaceuticals, textile, and bioethanol industries [<a href=\"#r-6\">6-10</a>]. Though, the sources of amylases are many such as plants, animals, bacteria and fungi, yet amylases of fungal origin are generally recognized as safe [<a href=\"#r-9\">9</a>]. That is also the reason, for a significant percentage of fungi are continuously screened for α-amylase or glucoamylase production with properties that are better suited to specific industrial applications [<a href=\"#r-11\">11-13</a>]. <em>A. flavus</em> has previously been recorded to be an active amylase enzyme producer [<a href=\"#r-13\">13-14</a>]. We have previously purified and sequences one noble α-amylase gene; and two nobles thermostable glucoamylase were also sequences and expressed from <em>A. favus </em>NSH9 [<a href=\"#r-15\">15- 17</a>]. One of its glucoamylase had starch binding domain (SBD) at C terminus and can degrade the raw starch granules [<a href=\"#r-17\">17</a>].<br />\r\nMicroorganisms can be easily controlled by means of genetic manipulation or other methods. These can be subject to strain development, mutations, genetic engineering, and other modifications that can increase the production of enzymes [<a href=\"#r-18\">18-19</a>]. The techniques used for strain improvement can be applied either separately or in different combinations [<a href=\"#r-20\">20</a>]. Random mutagenesis is widely used in the food industry for the classical strain development reasons [<a href=\"#r-21\">21</a>]. The mutagens are of chemical or physical in nature. Chemical mutagenesis includes the use of Ethidium bromides (EtBr), ethyl methyl sulphonate (EMS), nitrous acid, and N-methyl N’nitro-N nitroguanidine (NTG) [<a href=\"#r-22\">22</a>]. Physical mutagenesis encompasses UV rays, X-ray, and gamma rays. Among physical agents, UV treatment is commonly used in industries because it is very efficient and does not need any instruments [<a href=\"#r-22\">22</a>]. The natural ability of microorganisms to create amylase has indeed been enhanced by mutational therapy, where both chemical and UV radiation mutagenesis have been shown to increase amylase activity [<a href=\"#r-22\">22-24</a>]. Irradiation and chemical mutagens are mostly employed mutagens for mutations using random mutagenesis and some of them have considerably increased the characteristics of amylases in order to satisfy one or all of the 3 E’s i.e. Energy, Environment and Economy [<a href=\"#r-25\">25-27</a>]. The purpose of the study was to improve the production of amylases (α-amylase and two glucoamylases) from <em>A. flavus</em> NSH9 through random mutagenesis by using UV radiation and EtBr at different concentration at sublethal condition.</p>"
},
{
"section_number": 2,
"section_title": "MATERIALS AND METHODS",
"body": "<p><strong>Culture media for enzyme production</strong><br />\r\n<em>A. flavus</em> NSH9 has been used as an inoculum to produce amylase (α-amylase and two glucoamylases). So far as the enzyme production is concerned, active-growing fungal mycelium has been transferred from the potato dextrose agar ( PDA) plate to the minimum salt culture medium (MSM) containing (g / L): 4 g of yeast extract, 1 g (NH<sub>4</sub>)<sub>2</sub>SO<sub>4</sub>, 20 g raw sago starch, 3 g KH<sub>2</sub>PO<sub>4</sub> and 0.5 g MgSO<sub>4</sub>.7H<sub>2</sub>O [18, 26]. The pH was set to pH 5.0 prior to autoclaving. Two pieces (approximately 5 mm in diameter of each cutted piece) of a 7-day-old PDA fungal crop were used for fermentation in 250 mL of a conical flask containing 50 mL of MSM medium containing 2% (w / v) of raw sago starch. The incubation was conducted at room temperature for 5 days on a rotary shaker at 150 rpm. [<a href=\"#r-17\">17</a>, <a href=\"#r-28\">28</a>]. Details of sample preparation was discussed in the previous study [<a href=\"#r-28\">28</a>].</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Chemical and physical treatment for mutation</strong><br />\r\nFor chemical treatment in sublethal condition with EtBr at different concentration from 1µg/mL and 5µg/mL were added in the MNS culture media; and method is considered as treatment group one. The induction/fermentation procedure was also followed as discussed above.<br />\r\nOther chemical treatment with EtBr in PDA plate: In this method EtBr at concentration of 10µg/mL were added in potato dextrose agar (PDA) plate, and 100 µL of fungal spore suspension were spread onto the PDA plate. The process is considered as treatment group two. After that, the PDA plates were incubated for 7 days for growing mutated <em>A. flavus</em> NSH9. Two pieces of 7-day-old mutated fungal culture grown in PDA were then used for fermentation of 50 mL MSM medium. The incubation was carried out, as discussed above.<br />\r\nUV irradiation treatment was used as physical treatment for mutation and considered as treatment group three. Actively growing fungal mycelium was subjected to UV irradiation. First the fungal cultures grown actively for 5 days were suspended in 0.1M sodium acetate buffer (pH 5.0) and then were centrifuged for 10 min at 6000 rpm; this cell suspension was utilized for the mutation process. The mutation was induced by treatment with 10 mL of suspension which was aseptically pipetted into sterile flat-bottomed petri dishes. The sterile petri dishes containing the suspension were then exposed to UV light in a laminar airflow cabinet fitted with a germicidal lamp, according to Kumar et al. [<a href=\"#r-29\">29</a>]. The samples were exposed for 30 min. The mutated fungal cell suspensions (150 µL/plate) were spread onto PDA medium for 7 days. Afterward, two pieces of 7-day-old mutated fungal culture grown in PDA were used for fermentation of 50 mL MSM medium. The incubation was carried out, as discussed above.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Glucoamylase assay</strong><br />\r\nGlucoamylase activity was evaluated according to the technique used earlier by Karim et al. [<a href=\"#r-15\">15</a>] and released glucose was measured using 3, 5-dinitrosalicyclic acid (DNS) method [<a href=\"#r-30\">30</a>]. The absorbance measurements were performed at absorbance of 540 nm using a spectrophotometer. The enzyme activities were estimated using a calibration curve prepared with D-glucose as standard (Figure 1). One unit of glucoamylase activity is characterized as the amount of enzyme that releases 1 <em>µ </em>mole of glucose equivalent per minute from soluble starch under the test condition (at 55°C and pH 5.0) [<a href=\"#r-15\">15</a>].</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Raw starch degrading ability (RSDA) of glucoamylase</strong><br />\r\nThe degrading capacity of crude glucoamylase preparation to raw starch was determined by combining 0.5 mL of enzyme preparation with 0.5 mL of 1 percent (w / v) raw sago starch in 0.1 M sodium acetate buffer, pH 5.0. After 24 hours of incubation at 37 ° C with shaking at 150 rpm, the supernatant was collected for enzyme testing [<a href=\"#r-28\">28</a>]. The reaction mixture was assayed as described in above GA assay using standard curve of glucose (Figure 1). One unit of the RSDA was defined as the amount of enzyme required to release one µ mole of glucose per hour under the assay conditions.</p>\r\n\r\n<div id=\"figure1\">\r\n<figure class=\"image\"><img alt=\"\" height=\"290\" src=\"/media/article_images/2024/11/02/178-1597659391-Figure1.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 1. </strong>Glucose standard curve generated with DNS method at 540 nm. The concentration of glucose used in this study was ranging from 0.05 mg/mL to 0.25 mg/mL.</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Alpha amylase estimation</strong><br />\r\nα-amylase activity has been calculated using starch-iodine method according to Xiao et al. [<a href=\"#r-31\">31</a>] with minor modifications as discussed in previous study [<a href=\"#r-16\">16</a>]. A standard curve of the starch-iodine complex was prepared using a different amount of starch in 400 μL samples containing 50–400 μg of starch (<a href=\"#figure2\">Figure 2</a>), and the absorbance was measured at 580 nm. α-amylase activity unit (U) was defined in the starch-iodine assay as the disappearance in the assay reaction of an average of 1 mg of iodine binding starch material per min per mL.</p>\r\n\r\n<div id=\"figure2\">\r\n<figure class=\"image\"><img alt=\"\" height=\"333\" src=\"/media/article_images/2024/11/02/178-1597659391-Figure2.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 2. </strong>Absorbance of the starch-iodine complex standard curve generated with soluble starch at 580nm. The amount of starch used in the standard curve was ranging from 50 to 400 microgram. Calculated, Y = 10.0428 Absorbance (starch-iodine complex) at 580 nm for 1 mg of starch.</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>"
},
{
"section_number": 3,
"section_title": "RESULTS",
"body": "<p><strong>Effect on crude glucoamylase</strong><br />\r\nIn this study, UV radiation and EtBr at different concentrations in sublethal condition and in PDA plate were used to improve the production of GA, RSD-GA, and α-amylase. The crude GA production was comparatively higher in different treatment groups compared to control (without treatment) group (0.913 ± 0.09), and the production was significantly (<em>p</em> ≤0.05) different between the groups as shown by one-way ANOVA (<a href=\"#figure3\">Figure 3</a>). The highest GA activity (1.47 ± 0.087 U/mL/min) was observed in the culture media of the treatment group two with 10µg/ml EtBr in PDA plate method (<a href=\"#figure3\">Figure 3</a>), and the value was about 1.6 times higher compared to without treatment group. This value was significantly higher from all groups (treatment and control) except the treatment group with 5 µg/ml EtBr (1.387 ± 0.101) in sublethal condition (<em>p</em> ≤0.05, by Tukey test).</p>\r\n\r\n<div id=\"figure3\">\r\n<figure class=\"image\"><img alt=\"\" height=\"230\" src=\"/media/article_images/2024/11/02/178-1597659391-Figure3.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 3.</strong> Crude glucoamylase activity in different treatment by chemical and physical mutagens. The highest activity was recorded from mutated <em>A. flavus</em> NSH9 by EtBr in PDA plate (10µg/mL). Error bars show standard deviation among three independent observations.</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Effect on raw starch degrading glucoamylase</strong><br />\r\nLike GA, RSD-GA production also gradually expanded along with increasing concentration of EtBr at sublethal state (<a href=\"#figure4\">Figure 4</a>). As compared to control, all the treatment groups produced significantly higher RSD-GA (<em>p</em> ≤0.05). The maximum RSD-GA activity (0.839 ± 0.036 U/mL/Hour) was observed in the culture media of the treatment group two with 10µg/ml EtBr in PDA plate method (Figure 4). The highest value was about 1.5 times higher than control, which was remarkably close to treatment groups with UV and EtBr at 5µg/ml concentration (<a href=\"#figure4\">Figure 4</a>). Meanwhile, UV induced radiation also produced about 1.4 times more RSD-GA as compared to control (<a href=\"#figure4\">Figure 4</a>).</p>\r\n\r\n<div id=\"figure4\">\r\n<figure class=\"image\"><img alt=\"\" height=\"218\" src=\"/media/article_images/2024/11/02/178-1597659391-Figure4.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 4. </strong>Raw Starch Degrading glucoamylase (RSDG) activity in different treatment by chemical and physical mutagens. The highest activity was recorded from mutated NSH9 by EtBr in PDA plate (10µg/mL). Error bars show standard deviation among three independent observations.</figcaption>\r\n</figure>\r\n</div>\r\n\r\n<p style=\"text-align:justify\"> </p>\r\n\r\n<p style=\"text-align:justify\"><strong>Effect on α-amylase</strong><br />\r\nLike crude GA and RSD-GA, α-amylase production also differed significantly in various groups as found by one-way ANOVA (<em>p</em> ≤0.05). Compared to control, all the treatment groups produced significantly higher (<em>p</em> ≤0.05) α-amylase except the one with EtBr at low concentration (1µg/ml EtBr in sublethal condition).<br />\r\nThe highest α-amylase activity (0.162 ± 0.011 U/mL/min) was observed with 5µg/ml EtBr in sublethal condition (<a href=\"#figure5\">Figure 5</a>), and that was about 1.7 times higher than the control group. Meanwhile, UV treatment also produced about 1.63 times more α-amylase as compared to control (Figure 5). This highest value was significantly higher from only the control and treatment group with 1 µg/ml EtBr (0.108 ± 0.017) in sublethal condition (<em>p</em> ≤0.05, by Tukey test).</p>\r\n\r\n<div id=\"figure5\">\r\n<figure class=\"image\"><img alt=\"\" height=\"259\" src=\"/media/article_images/2024/11/02/178-1597659391-Figure5.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 5. </strong>α-amylase activity in different treatment by chemical and physical mutagens. The highest activity was recorded from <em>A. flavus</em> NSH9 by EtBr at sublethal condition in the culture (5 µg/mL). Error bars show standard deviation among three independent observations.</figcaption>\r\n</figure>\r\n</div>"
},
{
"section_number": 4,
"section_title": "DISCUSSION",
"body": "<p>Previous studies demonstrated that <em>A. flavus</em> NSH9 is a potential candidate for α-amylase, GA, and RSD-GA [<a href=\"#r-10\">10</a>, <a href=\"#r-15\">15-17</a>, <a href=\"#r-28\">28</a>]. So, it’s improvement through chemical and physical mutagens will be more industrial significance for amylases production [<a href=\"#r-19\">19</a>]. This study shows the impact of chemical and physical mutagens on the ability of amylase production from <em>A. flavus</em> NSH9. Like previous studies, this study exhibited a significant increase in amylases (all three enzymes) production by mutagens EtBr and UV rays [<a href=\"#r-25\">25-26</a>, <a href=\"#r-32\">32-35</a>]. The production of amylase (glucoamylase and amylase) by EtBr was higher compared to the UV radiation method in this study. A previous study observed that chemical mutagens (EtBr) are more capable of production glucoamylase from <em>Aspergillus niger</em> than physical mutagen (UV), which is similar to this study [<a href=\"#r-32\">32</a>]. They also reported that combination of both the EtBr and EMS can give the best result for glucoamylase production [<a href=\"#r-32\">32</a>]. About 1.5 times higher RSD-GA/RSDE activity was observed in this study with the EtBr treatment at 10µg/mL in PDA plate compared to wild strains, whereas a previous study recorded twofold higher RSDE from <em>Aspergillus</em> sp by combine treatments of γ-irradiation of Co<sup>60</sup>, UV and NTG [<a href=\"#r-34\">34</a>]. The higher amount of α-amylase from the mutated <em>A. flavus</em> NSH9 by EtBr at 5μg/mL in sublethal condition was found in the study, which is comparable to the previous research in sublethal cellulose production [<a href=\"#r-36\">36</a>]. Like the previous study, the chemically mutated strain produced a higher amount of α-amylase compare to UV mutated strains [<a href=\"#r-26\">26</a>].<br />\r\nThe research has few drawbacks, as the study used only one stage mutation technique, whereas the combination mutation method or the sequential mutation technique would have been better suited to the selection of potent mutants for the hyper-production of the desired enzyme. The research did not examine the nucleotide sequence of the intended mutated enzyme (amylases) to know the exact mutation point of the gene sequence. The study did not measure the toxic level of EtBr while using enzyme production at sublethal concentration.</p>"
},
{
"section_number": 5,
"section_title": "CONCLUSION",
"body": "<p>The higher production of both glucoamylases was recorded from mutated <em>A. flavus</em> NSH9 by EtBr in the PDA plate. Whereas the highest α-amylase production was recorded from the same fungi by EtBr at a concentration of 5µg/ml in the culture media. Although chemical treatment appears to have been more effective in improving the production of all amylases by fungal strain testing, UV radiation also increased the production of all enzymes. As the single mutation process of <em>A. flavus</em> NSH9 enhanced all three amylase enzymes, the strains could be used in the industrial development of amylase, and thus could be potential sources of starch processing.</p>"
},
{
"section_number": 6,
"section_title": "ACKNOWLEDGEMENT",
"body": "<p>None.</p>"
},
{
"section_number": 7,
"section_title": "AUTHOR CONTRIBUTIONS",
"body": "<p>KMRK proposed the original idea and reviewed the scientific contents described in the manuscript. KMRK and AR performed the experiments and analyzed the data. Both KMRK and AR wrote the initial draft and reviewed the finial manuscripts. The authors received no financial support for the research, authorship, and publication of this manuscript.</p>"
},
{
"section_number": 8,
"section_title": "CONFLICTS OF INTEREST",
"body": "<p>There is no conflict of interest among the authors.</p>"
}
],
"figures": [
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/11/02/178-1597659391-Figure1.jpg",
"caption": "Figure 1. Glucose standard curve generated with DNS method at 540 nm. The concentration of glucose used in this study was ranging from 0.05 mg/mL to 0.25 mg/mL.",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/11/02/178-1597659391-Figure2.jpg",
"caption": "Figure 2. Absorbance of the starch-iodine complex standard curve generated with soluble starch at 580nm. The amount of starch used in the standard curve was ranging from 50 to 400 microgram. Calculated, Y = 10.0428 Absorbance (starch-iodine complex) at 580 nm for 1 mg of starch.",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/11/02/178-1597659391-Figure3.jpg",
"caption": "Figure 3. Crude glucoamylase activity in different treatment by chemical and physical mutagens. The highest activity was recorded from mutated A. flavus NSH9 by EtBr in PDA plate (10µg/mL). Error bars show standard deviation among three independent observations.",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/11/02/178-1597659391-Figure4.jpg",
"caption": "Figure 4. Raw Starch Degrading glucoamylase (RSDG) activity in different treatment by chemical and physical mutagens. The highest activity was recorded from mutated NSH9 by EtBr in PDA plate (10µg/mL). Error bars show standard deviation among three independent observations.",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/11/02/178-1597659391-Figure5.jpg",
"caption": "Figure 5. α-amylase activity in different treatment by chemical and physical mutagens. The highest activity was recorded from A. flavus NSH9 by EtBr at sublethal condition in the culture (5 µg/mL). Error bars show standard deviation among three independent observations.",
"featured": false
}
],
"authors": [
{
"id": 533,
"affiliation": [
{
"affiliation": "Institute of Nutrition and Food Science, University of Dhaka, Dhaka-1000, Bangladesh"
}
],
"first_name": "Ashika",
"family_name": "Ruaida",
"email": null,
"author_order": 1,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 130
},
{
"id": 534,
"affiliation": [
{
"affiliation": "Institute of Nutrition and Food Science, University of Dhaka, Dhaka-1000, Bangladesh"
}
],
"first_name": "Kazi Muhammad Rezaul",
"family_name": "Karim",
"email": "rkarim98@gmail.com",
"author_order": 2,
"ORCID": null,
"corresponding": true,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "Kazi Muhammad Rezaul Karim, Associate Professor, Institute of\r\nNutrition and Food Science, University of Dhaka, Dhaka-1000,\r\nBangladesh, Email: rkarim98@gmail.com",
"article": 130
}
],
"views": 773,
"downloads": 102,
"references": [
{
"id": 4426,
"serial_number": 1,
"pmc": null,
"reference": "Rodriguez VB, Alameda EJ, Gallegos J, Requena AR, Lopez A. Enzymatic Hydrolysis of Soluble Starch with an alpha amylase from Bacillus licheniformis. Biotechnol Progr. 2006; 22: 718–22.",
"DOI": null,
"article": 130
},
{
"id": 4427,
"serial_number": 2,
"pmc": null,
"reference": "van der Maarel, MJEC, van der Veen B, Uitdehaag, JCM, Leemhuis H. and Dijkhuizen L. Properties and applications of starch-converting enzymes of the alpha-amylase family. J Biotechnol. 2002; 94: 137–55.",
"DOI": null,
"article": 130
},
{
"id": 4428,
"serial_number": 3,
"pmc": null,
"reference": "Leman P, Goesaert H, and Delcour JA. Residual amylopectin structures of amylase treated wheat slurries reflect amylase mode of action. Food Hydrocoll. 2009; 23: 153-164.",
"DOI": null,
"article": 130
},
{
"id": 4429,
"serial_number": 4,
"pmc": null,
"reference": "Sauer J, Sigurskjold BW, Christensen U, Frandsen TP, Mirgorodskaya E, Harrison M, Roepstorff P, and Svensson B. Glucoamylase: structure/function relationships, and protein engineering. Biochim Biophys Acta. 2000; 1543: 275-293.",
"DOI": null,
"article": 130
},
{
"id": 4430,
"serial_number": 5,
"pmc": null,
"reference": "Nguyen QD, Rezessy-Szabo JM, Claeyssens M, Stals I and Hoschke A. Purification and characterization of amylolytic enzymes from thermophilic fungus Thermomyces lanuginosus strain ATCC 34626. Enzyme Microb Technol. 2002; 31: 345-52.",
"DOI": null,
"article": 130
},
{
"id": 4431,
"serial_number": 6,
"pmc": null,
"reference": "Abdalwahab SA, Ibrahim SA and Dawood ES. Culture condition for the production of glucoamylase enzyme by different isolates of Aspergillus spp. Int Food Res J. 2012; 19: 1261-66.",
"DOI": null,
"article": 130
},
{
"id": 4432,
"serial_number": 7,
"pmc": null,
"reference": "Pandey A, Nigam P, Soccol VT, Singh D and Mohan R. Advances in microbial enzymes. Biotechnol Appl Biochem. 2000; 31: 135-152.",
"DOI": null,
"article": 130
},
{
"id": 4433,
"serial_number": 8,
"pmc": null,
"reference": "Kumar P and Satyanarayana T. Microbial glucoamylases: characteristics and applications. Crit Rev Biotechnol. 2009; 29: 225-255.",
"DOI": null,
"article": 130
},
{
"id": 4434,
"serial_number": 9,
"pmc": null,
"reference": "Gupta R, Gigras P, Mohapatra H, Goswami VK, Chauhan B. Microbial α-amylases: a biotechnological perspective. Process Biochem. 2003; 38: 1599–1616",
"DOI": null,
"article": 130
},
{
"id": 4435,
"serial_number": 10,
"pmc": null,
"reference": "Karim KMR and Tasnim T. (2018). Fungal glucoamylase production and characterization: A review. Rioresearch Communications. 2018;4: 591-605.",
"DOI": null,
"article": 130
},
{
"id": 4436,
"serial_number": 11,
"pmc": null,
"reference": "Negi S and Banerjee R. Optimization of extraction and purification of glucoamylase produced by Aspergillus awamori in solid-state fermentation. Biotechnol Bioprocess Eng. 2009; 14: 60-66.",
"DOI": null,
"article": 130
},
{
"id": 4437,
"serial_number": 12,
"pmc": null,
"reference": "Sanghvi GV, Koyani RD and Rajput KS. Isolation, optimization, and partial purification of amylase from Chrysosporium asperatum by submerged fermentation. J Microbiol Biotechnol. 2011; 21: 470–176.",
"DOI": null,
"article": 130
},
{
"id": 4438,
"serial_number": 13,
"pmc": null,
"reference": "Fadahunsi IF and Garuba OE. Amylase production by Aspergillus flavus associated with the bio-deterioration of starch-based fermented foods. N Y Sci J. 2012; 5: 13-18.",
"DOI": null,
"article": 130
},
{
"id": 4439,
"serial_number": 14,
"pmc": null,
"reference": "El-Abyad MS, Fawzeya A, El-Sayed A and Hafez M. Effects of culture conditions on amylase production by some soil fungi. Zbl Mikrob. 1992; 147: 23-34.",
"DOI": null,
"article": 130
},
{
"id": 4440,
"serial_number": 15,
"pmc": null,
"reference": "Karim KM R, Husaini A, Hossain MA, Sing NN, Mohd Sinang F, Hussain MHM and Roslan HA. Heterologous, Expression, and Characterization of Thermostable Glucoamylase Derived from Aspergillus flavus NSH9 in Pichia pastoris. BioMed Res Int. 2016; 5962028.",
"DOI": null,
"article": 130
},
{
"id": 4441,
"serial_number": 16,
"pmc": null,
"reference": "Karim KMR, Husaini A, Sing NN, Mohd Sinang F, Roslan HA, Hussain H. Purification of an alpha amylase from Aspergillus flavus NSH9 and molecular characterization of its nucleotide gene sequence. 3 Biotech. 2018; 8. 204.",
"DOI": null,
"article": 130
},
{
"id": 4442,
"serial_number": 17,
"pmc": null,
"reference": "Karim KMR, Husaini A, Sing NN, Tasnim T, Sinang FM, Hussain H, Hossain MA and Roslan H. Characterization and Expression in Pichia pastoris of a Raw Starch Degrading Glucoamylase (GA2) derived from Aspergillus flavus NSH9. Protein Expr Purif. 2019;164: 105462.",
"DOI": null,
"article": 130
},
{
"id": 4443,
"serial_number": 18,
"pmc": null,
"reference": "Ajita S and Thirupathihalli PKM. α-Amylase Production and Applications: A Review. J Appl Environ Microbiol. 2014; 2: 166-175.",
"DOI": null,
"article": 130
},
{
"id": 4444,
"serial_number": 19,
"pmc": null,
"reference": "Makino T, Skretas G, Georgiou G. Strain engineering for improved expression of recombinant proteins in bacteria. Microb Cell Fact. 2011; 10:32.",
"DOI": null,
"article": 130
},
{
"id": 4445,
"serial_number": 20,
"pmc": null,
"reference": "Nevalainen K. Strain improvement in filamentous fungi-an overview. Appl Microbiol Biotechnol. 2001; 1: 289–304.",
"DOI": null,
"article": 130
},
{
"id": 4446,
"serial_number": 21,
"pmc": null,
"reference": "Margolles A, Sanchez B. Selection of a Bifidobacterium animalis subsp. lactis strain with a decreased ability to produce acetic acid. Appl Environ Microbiol. 2012; 78: 3338–42.",
"DOI": null,
"article": 130
},
{
"id": 4447,
"serial_number": 22,
"pmc": null,
"reference": "Pathak SS, Sandhu SS, Rajak RC. 2015. Mutation studies on fungal glucoamylase: a review. Int J Pharm Biol Sci. 2015; 5: 297-308",
"DOI": null,
"article": 130
},
{
"id": 4448,
"serial_number": 23,
"pmc": null,
"reference": "EL-Bondkly AM and Keera AA. UV-and EMS-induced mutations affecting synthesis of alkaloids and lipase in Penicillium roquefortii. Arab J Biotechnol. 2007; 10: 241–248.",
"DOI": null,
"article": 130
},
{
"id": 4449,
"serial_number": 24,
"pmc": null,
"reference": "Vu VH, Pham TA, and Kim K. Improvement of fungal cellulase production by Mutation and optimization of solid state fermentation. Mycobiol. 2011; 39: 20–25.",
"DOI": null,
"article": 130
},
{
"id": 4450,
"serial_number": 25,
"pmc": null,
"reference": "Aleem B, Rashid MH, Zeb N. and et al. Random mutagenesis of super Koji (Aspergillus oryzae): improvement in production and thermal stability of α-amylases for maltose syrup production. BMC Microbiol. 2018; 18: 200.",
"DOI": null,
"article": 130
},
{
"id": 4451,
"serial_number": 26,
"pmc": null,
"reference": "Singh S, Singh S and Mangla J. Physical and Chemical Mutation for Enhanced Alpha-Amylase Production by Aspergillus fumigatus NTCC1222 under Solid State Fermentation Conditions Using Agri-Residue Waste. J Pharm Nutr Sci. 2016; 6: 22-26.",
"DOI": null,
"article": 130
},
{
"id": 4452,
"serial_number": 27,
"pmc": null,
"reference": "Singh S, Dutt D, Tyagi CH, Upadhyaya JS. Bio-conventional bleaching of wheat straw soda-AQ pulp with crude xylanases from SH-1 NTCC-1163 and SH-2 NTCC-1164 strains of Coprinellus disseminatus to mitigate AOX generation. New Biotechnol. 2010; 28: 47-57.",
"DOI": null,
"article": 130
},
{
"id": 4453,
"serial_number": 28,
"pmc": null,
"reference": "Karim KMR, Husaini A and Tasnim T. Production and Characterization of Crude Glucoamylase from Newely isolated Aspergillus flavus NSH9 in Liquid Culture. Am J Biochem Mol Biol. 2017; 7: 118-26.",
"DOI": null,
"article": 130
},
{
"id": 4454,
"serial_number": 29,
"pmc": null,
"reference": "Kumar GS, Chandra MS, Sumanth M, Vishnupriya A, Reddy BR, and Choi YL. Cellulolytic enzymes production from submerged fermentation of different substrates by newly isolated Bacillus spp. FME. K S J Appl Biol Chem. 2009; 52: 17-21.",
"DOI": null,
"article": 130
},
{
"id": 4455,
"serial_number": 30,
"pmc": null,
"reference": "Miller GL. Use of dinitrosalicylic acid reagent for determination of reducing sugars. Anal Chem. 1959; 31: 426–428.",
"DOI": null,
"article": 130
},
{
"id": 4456,
"serial_number": 31,
"pmc": null,
"reference": "Xiao Z, Storms R and Tsang A. (2006). A quantitative starch-iodine method for measuring alpha-amylase and glucoamylase activities. Anal Biochem. 2006; 351: 146-48.",
"DOI": null,
"article": 130
},
{
"id": 4457,
"serial_number": 32,
"pmc": null,
"reference": "Raju EVN, Divakar G, Swetha C, Geetha J, Satish P. Strain improvement of Aspergillus niger for glucoamylase by physical and chemical mutagens. Int Res J Pharm App Sci. 2012; 2: 79-91.",
"DOI": null,
"article": 130
},
{
"id": 4458,
"serial_number": 33,
"pmc": null,
"reference": "Vu VH, Pham TM, Kim K. Improvement of a fungal strain by repeated and sequential mutagenesis and optimization of solid state fermentation for the hyperproduction of raw starch digesting enzyme. J Mirobiol Biotechnol. 2010; 20: 718-26.",
"DOI": null,
"article": 130
},
{
"id": 4459,
"serial_number": 34,
"pmc": null,
"reference": "Vu VH and Keun K. Hyper-production of raw starch-digesting enzyme by mutant fungal strain and optimization of solid byproducts. J Viet Env. 2012; 3: 66-70.",
"DOI": null,
"article": 130
},
{
"id": 4460,
"serial_number": 35,
"pmc": null,
"reference": "Oluwatoyin F, Olukunle and Oluwadamilola E, Ajayi. 2018. Screening Wild and Mutant Strains of Aspergillus flavus and Aspergillus niger Isolated from Plantain Stalks for Amylase Production. Jordan J Biol Sci. 2018; 11: 557 – 62.",
"DOI": null,
"article": 130
},
{
"id": 4461,
"serial_number": 36,
"pmc": null,
"reference": "Chand P, Aruna A, Maqsood AM and Rao LV. Novel mutation method for increased cellulase production. J. Appl. Microbiol. 2005; 98: 318-23.",
"DOI": null,
"article": 130
}
]
},
{
"id": 120,
"slug": "178-1597401409-comparative-molecular-analysis-of-contemporary-isolates-of-duck-plague-virus-from-haor-areas-of-bangladesh",
"featured": false,
"slider": false,
"issue": "Vol4 Issue1",
"type": "short_communication",
"manuscript_id": "178-1597401409",
"recieved": "2020-08-11",
"revised": null,
"accepted": "2020-10-17",
"published": "2020-10-24",
"pdf_file": "https://jabet.bsmiab.org/media/pdf_file/2023/16/178-1597401409.pdf",
"title": "Comparative molecular analysis of contemporary isolates of duck plague virus from haor areas of Bangladesh",
"abstract": "<p>Duck plague (DP) is one of the most important viral diseases which affects the duck population across the globe including Bangladesh. The present work was conducted to detect DP virus (DPV) from <em>haor</em> areas using a molecular-based approach and compared with the contemporary isolate through molecular and phylogenetic analysis. For this purpose, 38 individual samples were collected from the Netrokona (n=20) district of the Mymensingh division and Kishoreganj (n=18) district of the Dhaka division. The identification of DVP was carried out by polymerase chain reaction (PCR) targeting DPV specific <em>DNA polymerase </em>genes followed by sequencing. PCR positive viral samples were used to propagate into 11-13 days old embryonated duck eggs through chorio-allantoic membrane (CAM) route for virus isolation. DPV were then propagated into duck embryo fibroblast (DEF) monolayer cell culture and confirmed by PCR. Among the 38 samples, 27 isolates were confirmed as DPV with the PCR amplicon size of 446 bp. Pathogenicity tests through the inoculation into day-old ducklings confirmed pathogenic strain. The PCR products of the isolated DPV specific DNA polymerase gene were sequenced commercially and submitted to GenBank (GenBank Accession No. KX768734.1). The sequence showed resemblance to isolates previously reported in India (GenBank Accession No. KX511893.1, KJ451479.1, KM012009.1), and China (GenBank Accession No. EF643559.1). Sequencing data also revealed nucleotide differences between Anatid herpes 1_BAU_DMH (previous report from our laboratory) and the present isolates. Further characterization, such as nucleotide and amino acid sequencing, would help to understand the strains along with its epidemiology.</p>",
"journal_reference": "J Adv Biotechnol Exp Ther. 2021; 4(1): 44-52.",
"academic_editor": "Md Jamal Uddin, PhD; Ewha Womans University, Seoul, South Korea",
"cite_info": "Khan MT, Pavel MTR, et al. Comparative molecular analysis of contemporary isolates of duck plague virus from haor areas of Bangladesh. J Adv Biotechnol Exp Ther. 2021; 4(1): 44-52.",
"keywords": [
"PCR",
"DNA polymerase gene",
"Duck plague virus",
"Sequencing"
],
"DOI": "10.5455/jabet.2021.d105",
"sections": [
{
"section_number": 1,
"section_title": "INTRODUCTION",
"body": "<p>Duck plague (DP; synonym: duck viral enteritis), is the highly infectious and contagious disease of ducks, causing mortality of 60-70% [<a href=\"#r-1\">1</a>] in Bangladesh. It is considered as one of the potential threats to commercially reared, domestic and wild waterfowl [<a href=\"#r-2\">2</a>] throughout the world as it affects all age groups of ducks. DP is caused by duck herpesvirus 1 (anatid herpesvirus 1) under the genus <em>Mardivirus</em> of the family Herpesviridae and subfamily <em>Alphaherpesvirinae </em>[<a href=\"#r-3\">3</a>]. Like other herpesviruses, this virus has a linear double-stranded DNA genome of 119×10<sup>6</sup> Daltons and comprised of approximately 158,091 base pairs [<a href=\"#r-4\">4</a>]. 65 out of 67 genes are found as coding genes and three genes are unique to AHV-1 and the genome is made of unique long (UL), unique short (US), unique short internal repeat (IRS), and unique short terminal repeat (TRS) regions (UL-IRS-US-TRS in order) [<a href=\"#r-4\">4</a>].<br />\r\nDirect or indirect contact with infected ducks or contaminated environment (that is contaminated with feces or oral and nasal secretions from an infected bird), respectively, can be the main mode of transmission of duck plague virus (DPV) [<a href=\"#r-5\">5</a>]. Migratory waterfowls act as asymptomatic carriers of disease. In natural infection, incubation period varies from 3 to 7 days [<a href=\"#r-6\">6</a>]. In most cases, the first notification is abrupt and consistently increasing flock mortality and infected ducks may die in good flesh though no symptoms are detectable. Affected birds show various general symptoms like other diseases, but prolapse of penis in dead mature male breeders and a marked reduction in egg production in female are remarkable, and mortality may reach up to 100% [<a href=\"#r-7\">7</a>]. Diphtheroid plaques on the eyelids and the mucosae of the respiratory system and gastrointestinal system usually result in ophthalmic signs and refusal to water [<a href=\"#r-6\">6</a>].<br />\r\nIn the context of Bangladesh, the first report and confirmation of DPV had been made in the year 1982 [<a href=\"#r-1\">1</a>], and later isolated and characterized by other investigators as well [<a href=\"#r-8\">8-10</a>]. Hossain <em>et al</em>. [<a href=\"#r-11\">11</a>] evaluated the immunogenicity of experimentally developed DPV vaccine from local isolates. DPV can be propagated and detected in duckling or adult duck [<a href=\"#r-4\">4</a>], 9-12 days old embryonated duck eggs through chorio-allantoic membrane (CAM) route [<a href=\"#r-9\">9,12</a>], avian fibroblast cell [<a href=\"#r-13\">13</a>], kidney cell, liver cell etc. This virus can also be identified by passive haemagglutination (PHA) test [<a href=\"#r-11\">11,14</a>]; neutralization test [<a href=\"#r-5\">15</a>] or by polymerase chain reaction (PCR) with specific primer [<a href=\"#r-10\">10</a>] and PCR is nearly 20 times more sensitive than tissue culture, and more reliable and accurate than traditional virus isolation and serologic identification methods used for detecting duck plague virus [<a href=\"#r-16\">16</a>].<br />\r\nIn Bangladesh, the total duck population is 54.016 million and ranked second in poultry, next to chicken [<a href=\"#r-17\">17</a>]. Ducks are found throughout the country, especially in the north-east wetland ecosystem (<em>haor</em>) [<a href=\"#r-18\">18</a>] and with more than 24% of the country’s total duck population found in this <em>haor</em> region [<a href=\"#r-19\">19</a>]. In Netrokona and Kishoreganj districts, a higher number of duck populations is recorded than the other districts and provides self-employment for the landless and marginal small farmers in this region [<a href=\"#r-18\">18</a>]. As DP is the most important disease of ducks, proper detection, isolation including molecular detection and analysis of the virus are of great importance. The molecular researches on duck plague virus are lag behind the other members of the herpesviridae family and even before this decade, mainly focused on epidemiology and prevention of the disease [<a href=\"#r-20\">20</a>]. The genomic organization and nucleotide sequencing might be helpful for further study on duck plague virus [<a href=\"#r-4\">4</a>]. Hence, UL30 gene (encoding DNA polymerase protein) based PCR detection, nucleotide sequencing and phylogenetic analysis was previously reported from our laboratory [<a href=\"#r-10\">10</a>]. Subsequently, this present research work was undertaken as a follow up and comparative analysis to understand the strains and its epidemiology. Therefore, the present research was conducted for the isolation, identification, comparative molecular and phylogenetic analysis of DPV from <em>haor</em> areas in Bangladesh.</p>"
},
{
"section_number": 2,
"section_title": "MATERIALS AND METHODS",
"body": "<p><strong>Study area and period</strong><br />\r\nThe experimental work was conducted at the Virology Laboratory, Department of Microbiology and Hygiene, BAU, Mymensingh, Bangladesh, from July 2015 to August 2016. A total of 38 randomly selected ducks (moribund and dead) from Sadar upazila of Netrokona district (n=20) of Mymensingh division and Tarail upazila (n=18) of Kishoreganj district of Dhaka division of Bangladesh. The map of the sampling locations is shown in the <a href=\"#figure1\">Figure 1</a>.</p>\r\n\r\n<div id=\"figure1\">\r\n<figure class=\"image\"><img alt=\"\" height=\"260\" src=\"/media/article_images/2024/58/03/178-1597401409-Figure1.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 1.</strong> Maps of sampling locations. Dash circle indicating sadar upazila of Netrokona district and solid circle indicating Tarail upazila of Kishoreganj district. (Source: <a href=\"http://www.mapsofbangladesh.com/\">http://www.mapsofbangladesh.com/</a>)</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Sampling</strong><br />\r\nDifferent visceral organs, such as the esophagus, liver, intestine, and proventriculus, were collected aseptically during the post-mortem examination of 38 suspected DP affected ducks. The collected samples were kept separately in a sterile plastic container with proper labeling. Maintaining proper cool chain, the samples were then transported to the departmental Virology Laboratory, and few were processed immediately, and the rest were stored at ‑20°C until further analysis.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Preparation of inocula</strong><br />\r\nThe samples were processed by grinding, and PBS was added simultaneously to make a 10% suspension. Then centrifugation of the preliminary suspension was done at 4500 rpm for 10 min [<a href=\"#r-2\">2</a>] and the supernatant fluid was collected and then treated with antibiotics (Gentamycin, 100μg/mL of suspension). For the sterility test, antibiotic-treated inocula were streaked separately with a sterile inoculating loop over the nutrient agar media and fresh blood agar media [<a href=\"#r-11\">11</a>]. and the plate was incubated at 37<sup>0</sup>C for overnight.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Propagation of virus in embryonated duck eggs</strong><br />\r\nFor this purpose, the prepared sterile inocula were inoculated into 11-13 days old embryonated duck eggs (EDE) through CAM route [<a href=\"#r-2\">2</a>, <a href=\"#r-9\">9</a>], and at 6-8 days post-infection (PI), all dead and live EDEs was chilled overnight, and allantoic fluid and CAM were collected.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Propagation of the virus in duck embryo fibroblast cell culture</strong><br />\r\nDuck embryo fibroblast cell culture was prepared according to the methods described by OIE manual [<a href=\"#r-2\">2</a>] and Hossain et al [<a href=\"#r-11\">11</a>]. Briefly, embryonated duck eggs (11-13 days old) were collected and soaked with 70% ethanol. The embryos were taken out and placed on a petri dish, followed by chopping of embryos excluding extremities and viscera. Chopped embryos were then taken into a conical flask where 0.25% trypsin was added for 100 mg tissues for cold trypsinization (4<sup>0</sup>C for 18 hours), followed by the addition of 5% Minimum Essential Media (MEM) for minimization of effect of trypsin. Then the suspension was transferred into cell culture flasks and was incubated at 37<sup>0</sup>C temperature and periodically observed for cell growth. Cell culture flasks containing 75-80% confluent growth of cells were taken for virus propagation. The remaining media of the cell culture flask was discarded. Cells were washed two times with PBS. About 200 μl virus inoculum was spread over monolayer duck embryo fibroblast cells. Then the cell culture flask was incubated at 37<sup>0</sup>C for 60 min in an orbital shaker incubator to allow the virus to infect the cell. After incubation, 5% calf serum containing maintenance medium was added and incubated at 37<sup>0</sup>C temperature. A cell culture flask was observed daily under an inverted microscope for the development of cytopathic effect (CPE).</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Propagation of virus into ducklings</strong><br />\r\nThe duck plague virus was propagated into ducklings for pathogenicity test, according to the method described by Ahmad et al [<a href=\"#r-10\">10</a>] and for this purpose, ethical approval was taken [Ethical Approval no. AWEEC/BAU/2017(09)]. For pathogenicity tests, about 500 µl (10<sup>5.7</sup> dELD<sub>50</sub>/mL) DPV suspension (CAM suspension) was inoculated in ducklings through the intramuscular route and observed for 6-8 days. It was expected that all the inoculated ducklings would be affected and would show clinical signs after 6 days PI.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>DNA extraction, PCR and electrophoresis</strong><br />\r\nDNA extraction kit Wizard<sup>® </sup>Genomic DNA Purification Kit (Promega, Madison, Wisconsin, United States) was used to extract DNA following the manufacturer’s instructions. The desired DNA segments of targeting the <em>DNA polymerase </em>gene of DPV (expected amplicon size of 446 bp) was amplified by PCR using the primers (<a href=\"#Table-1\">Table 1</a>) as described by Wu et al<em>.</em> [<a href=\"#r-15\">15</a>]. The reaction mixture (total volume 25 µL) was comprised of nuclease-free water (7.5 µL), 2x PCR master mixture (12.5 µL; Promega, Madison, Wisconsin, United States), each forward and reverse primers (each 1 µL), and extracted DPV DNA template (3 µL). The thermal profile was set at initial denaturation at 94°C for 2 min; 35 cycles 94°C for 1 min, 56°C for 1 min, 72°C for 2 min, and, a final extension at 72°C for 7 min. For agarose gel electrophoresis of the PCR products, 2% agarose gel was used and PCR products (5 µL), including the DEV positive and native control, were mixed with 6X loading dye (Promega, USA; 1 µL) and loaded to the appropriate well. Agarose gel electrophoresis was accomplished following the method of Wu et al. [<a href=\"#r-15\">15</a>].</p>\r\n\r\n<div id=\"Table-1\">\r\n<p><a href=\"https://jabet.bsmiab.org/table/178-1597401409-table1/\">Table-1</a><strong>Table 1. </strong>Primers for PCR.</p>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Sequencing and analysis</strong><br />\r\nFor further molecular characterization, the PCR product of <em>DNA polymerase</em> gene (446-bp amplicon size) of DPV was used for partial sequencing. The PCR product was purified using Wizard<sup>® </sup>SV Gel and PCR Clean-Up system (Promega, Madison, Wisconsin, United States) according to their Quick protocol. Among the 27 positive isolates, two samples were sequenced, followed by analysis using BioEdit 7.0.5 Version [<a href=\"#r-21\">21</a>] and ClustalX 2.0 [<a href=\"#r-22\">22</a>] softwares. The finally obtained sequence was submitted to the GenBank. Then, the National Center for Biotechnology Information (NCBI) database (http://www.ncbi.nlm.nih.gov/) was used to analyze the sequencing results, and comparison between the results of sequence analyses and a few other published sequences was made. For the construction of the Phylogenetic tree, the genome sequences of total 24 published sequences were aligned using CLC Sequence Viewer 8.0 software and finally, the phylogenetic analysis was accomplished through CLC Sequence Viewer 8.0 based on Neighbor-Joining method (<a href=\"#figure7\">Figure 7</a>).</p>"
},
{
"section_number": 3,
"section_title": "RESULTS",
"body": "<p><strong>Isolation of duck plague virus</strong><br />\r\nDPV was isolated from 27 samples out of 38, i.e., the overall isolation rate was 71.05% and district wise isolation rate were 80% (16/20) and 61.11% (11/18) in Netrokona and Kishoreganj district, respectively.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Propagation in embryonated duck eggs</strong><br />\r\nThe allantoic fluid and the CAM were collected from the 11-13 days-old duck embryo which was inoculated with pure DPV suspension through CAM route. Embryo mortality in this study started from the 6 days PI and observed up to 8 days PI, and all the dead and live embryos were chilled at 4°C. Subcutaneous hemorrhages, thickened, and hemorrhagic CAM was found in dead embryos (<a href=\"#figure2\">Figure 2a, b</a>, and <a href=\"#figure2\">c</a>). Slide HA using the allantoic fluid revealed negative results (<a href=\"#figure2\">Figure 2d</a>).</p>\r\n\r\n<div id=\"figure2\">\r\n<figure class=\"image\"><img alt=\"\" height=\"209\" src=\"/media/article_images/2024/58/03/178-1597401409-Figure2.jpg\" width=\"323\" />\r\n<figcaption><strong>Figure 2.</strong> Results of propagation of present isolates of DPV in embryonated duck eggs. (a) normal embryo remained intact and CAM (arrow); (b) hemorrhagic thickened CAM and (c) petechial hemorrhages over the embryo’s body with no feather in DPV inoculated embryo; (d) Slide hemagglutination test mixing allantoic fluid from DPV infected embryo and 2% cRBC (chicken RBC) showed no hemagglutination.</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Cytopathic effects in duck embryo fibroblast cell culture</strong><br />\r\nAfter the adaptation of the virus in fibroblast cells, CPE was observed as enlarged, rounded, clumped, degenerated, and necrosed of fibroblast cells and giant cell formation (<a href=\"#figure3\">Figure 3</a>). Flasks with maximum CPE were frozen at -20<sup>0</sup>C or -80<sup>0</sup>C.</p>\r\n\r\n<div id=\"figure3\">\r\n<figure class=\"image\"><img alt=\"\" height=\"207\" src=\"/media/article_images/2024/58/03/178-1597401409-Figure3.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 3. </strong>Results of propagation of the present isolates of duck plague virus into duck embryo fibroblast cell culture. (a) Control: Growth pattern of duck embryo fibroblast cells (400X magnification), kept as control, showing confluent growth of star like fibroblast cells. (b) Infected: Observation CPE of duck embryo fibroblast cells by duck plague virus showing round, necrotic, degeneration of cells, clumping of cell and giant cell formation of fibroblast cells under green light (400X magnification)(arrow: clumping of cell; dash circle: giant cell; solid circle: round cell).</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Pathogenicity test in day-old duckling</strong><br />\r\nAll the inoculated ducklings were affected within 6 days of post-infection showing clinical signs. Ducklings were found unable to stand with head down and drooped out-stretched wings and other symptoms, such as weakness, depression, off food, ataxia, and diarrhea were also found. Nervous sings began to rise as tremors of head, body, and neck, and finally death occurred (Figure 4a). Unclotted blood in the body cavities, pinpoint hemorrhages in the pale liver, and hemorrhagic annular band in the intestine were found on postmortem examination (<a href=\"#figure4\">Figure 4b, c</a>, and <a href=\"#figure4\">d</a>). For the re-isolation of the virus, visceral organs were processed and extracted for DNA and finally confirmed by PCR.</p>\r\n\r\n<div id=\"figure4\">\r\n<figure class=\"image\"><img alt=\"\" height=\"396\" src=\"/media/article_images/2024/58/03/178-1597401409-Figure4.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 4. </strong>Results of pathogenicity test for the duck plague virus isolates. (a) Dead duckling after 6 days of post inoculation of DP virus; postmortem examination of ducklings showing (b) unclotted free blood in the body cavities, (c) petechial hemorrhage in the pale liver, (d) hemorrhagic annular band in intestine.</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Detection of DPV by PCR</strong><br />\r\nThe extracted DNA samples from 38 field isolates were used to amplify DNA polymerase genes by PCR and 27 samples were found positive for DPV. All the PCR positive products showed the expected amplicon size of 446-bp (<a href=\"#figure5\">Figure 5</a>).</p>\r\n\r\n<div id=\"figure5\">\r\n<figure class=\"image\"><img alt=\"\" height=\"174\" src=\"/media/article_images/2024/58/03/178-1597401409-Figure5.jpg\" width=\"320\" />\r\n<figcaption><strong>Figure 5.</strong> PCR amplification products of duck plague virus with DNA polymerase gene specific primer. Lane M: 100 bp ladder; Lane 1: positive control; Lane 2-6: positive samples of duck plague virus; Lane 7: negative control.</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Sequencing and phylogenetic analysis</strong><br />\r\nThe partially sequenced data were submitted to the GenBank (GenBank Accession No. KX768734.1) and the isolate was mentioned as BAU_DP_1T. Comparative analysis of DNA polymerase gene of DPV with previously reported isolates were performed and presented in <a href=\"#figure6\">Figure 6</a>. The sequenced data were used to construct a phylogenetic tree (<a href=\"#figure7\">Figure 7</a>). The tree revealed that the present isolates share the common ancestral origin with the isolates and strains found in Bangladesh, India, and China. The GenBank accession number and percent identities of the sequences of 24 isolates/strains were represented in the <a href=\"#Table-2\">Table 2</a>.</p>\r\n\r\n<div id=\"figure6\">\r\n<figure class=\"image\"><img alt=\"\" height=\"224\" src=\"/media/article_images/2024/58/03/178-1597401409-Figure6.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 6. </strong>Comparative analysis of the nucleic acid sequenced data of the present isolates and Ahamed et al 2015 [10]. The nucleotide variation was found in the 374-379 position. The ‘solid box’ indicated the difference region and the ‘dash box’ indicated the nucleotide sequence which was not reported in the previous study from our laboratory (i.e., Ahamed et al., 2015, reported 378 bp out of 446 bp amplicon).</figcaption>\r\n</figure>\r\n</div>\r\n\r\n<div id=\"figure7\">\r\n<figure class=\"image\"><img alt=\"\" height=\"242\" src=\"/media/article_images/2024/58/03/178-1597401409-Figure7.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 7. </strong>Phylogenetic tree showing relationship of DPV isolates. The tree was constructed through the neighbor joining method and nucleotide distances were measures through Jukes-Cantor method using CLC Sequence Viewer 8.0 software. Numbers on the nodes indicate bootstrap percentage values (calculated using 100 replicates). Red color represents this study isolates.</figcaption>\r\n</figure>\r\n</div>\r\n\r\n<div id=\"Table-2\">\r\n<p><a href=\"https://jabet.bsmiab.org/table/178-1597401409-table2/\">Table-2</a><strong>Table 2. </strong>List of isolates of DPV for homology study of fusion gene sequences.</p>\r\n\r\n<p> </p>\r\n</div>"
},
{
"section_number": 4,
"section_title": "DISCUSSION",
"body": "<p>Duck plague is considered the main obstacle in duck rearing and affects severely the large-scale duck farming in the <em>haor</em> and coastal areas of Bangladesh. The overall isolation rate (71.05%) of this study is in close agreement with the results reported by Hansen et al. [<a href=\"#r-23\">23</a>] and Ahamed et al<em>.</em> [<a href=\"#r-10\">10</a>]. In a previously published report from our laboratory, the overall prevalence was shown there as 66.67% [<a href=\"#r-24\">24</a>] where the DP prevalence was 71.42% and 66.67% in Netrokona and Sunamganj district, respectively, whereas, in this study, that values were 80% and 61.11% in Netrokona and Kishoreganj district, respectively.<br />\r\nThe cultural properties of the present isolates were observed both in embryonated duck eggs and cell culture system, and pathogenicity was indicated by inoculating in ducklings. The lesions found in this study (i.e., subcutaneous haemorrhage, thickened and hemorrhagic CAM) at 6 to 8 days PI, were consistent with the findings of Akter et al. [<a href=\"#r-9\">9</a>], El-Samadony et al. [<a href=\"#r-12\">12</a>] and Ahamed et al. [<a href=\"#r-10\">10</a>]. As virus titre was highest in CAM, the CAM route is considered as the most sensitive way in the indicator host system for DPV propagation. In case of duck embryo fibroblast cell culture, the findings were supported by Nguyen et al. [<a href=\"#r-25\">25</a>] and Aravind et al. [<a href=\"#r-26\">26</a>] where the authors also observed similar CPE caused by DP virus in Duck embryo fibroblast cells. The pathogenicity test of the present isolates revealed that the isolates are pathogenic, as all the ducklings showed typical clinical signs within 6 days of post-infection. The results of the pathogenicity test is supported by the findings of El-Samadony et al. [<a href=\"#r-12\">12</a>].<br />\r\nThe PCR result of the present study was consistent and in agreement with the published reports of Wu et al. [<a href=\"#r-15\">15</a>], Ahamed et al. [<a href=\"#r-10\">10</a>], and OIE manual [<a href=\"#r-2\">2</a>]. The use of the polymerase gene, for the PCR confirmation of DPV, is reported by various researchers from different countries of the world [<a href=\"#r-27\">27</a>, <a href=\"#r-28\">28</a>].<br />\r\nPhylogenetic tree demonstrated that the isolate of duck plague virus (BAU_DP_1T) in this study was almost similar with the nucleic acid sequenced data found from GenBank. The GenBank Accession No. with KJ549663.1|: Anatidherpesvirus_1 stra CV, AF064639.1|:77-448 Anatid herpesvirus 1 DNA polymerase gene, EF643559.1|:635-932 Duck enteritis virus UL31 protein (UL31) gene, JQ647509.1|:59034-59405 Anatid herpesvirus 1 strain CHv and EU082088.2|:55521-55892 Duck enteritis virus strain VAC which has been reported as originated in China and was responsible for using duck plague in domestic ducks and other waterfowls. The phylogenetic tree also showed similarities with some other nucleic acid sequenced data from GenBank (<a href=\"#figure7\">Figure 7</a>). We assume that the present isolate could be originated from the West Bengal and Kerala regions of India (KX511893.1: DEV/India/IVRI-2016; KM012009.1: isolate CDIO-duck01, Kerala, India 2015; KJ451479.1: DEV/Atpara/13, West Bengal, India 2014).<br />\r\nComparative sequencing result with the previously published one [<a href=\"#r-10\">10</a>], showed differences in few nucleotide base pairs (nucleotide position 374 to 379). Moreover, the previous authors reported only 378 bp among 446 bp PCR product. Therefore, further molecular analysis, such as nucleotide sequencing, amino acid sequencing etc. are of great significance.</p>"
},
{
"section_number": 5,
"section_title": "CONCLUSION",
"body": "<p>Present research work revealed that the overall isolation rate was 71.05% (27 DPV positive isolates out of 38 suspected samples). PCR with <em>DNA polymerase</em> gene-specific primers and sequencing confirmed the isolates as DPV. The pathogenicity test also revealed that the field isolates were to be pathogenic. Nucleic acid sequencing of PCR products of 446-bp and phylogenetic tree showed that this isolate of duck plague virus (BAU_DP_1T) was highly similar with the isolates of DPV strain which were reported from India and China.</p>"
},
{
"section_number": 6,
"section_title": "ACKNOWLEDGMENTS",
"body": "<p>None.</p>"
},
{
"section_number": 7,
"section_title": "CONFLICTS OF INTEREST",
"body": "<p>There is no conflict of interest among the authors.</p>"
},
{
"section_number": 8,
"section_title": "AUTHOR CONTRIBUTIONS",
"body": "<p>MTK conducted the main research and both MTK and MBR was involved in conception and design of the experiment. MTRP and AK contributed to perform the experiment. TF helped in giving outbreaks information and sample collection. MAHS, MTR and KHMNHN were involved in sequence analysis. MTH and MPS contributed to prepare the manuscript and MBR finally made the approval of the version to be published. No funding was provided for this research work.</p>"
}
],
"figures": [
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/58/03/178-1597401409-Figure1.jpg",
"caption": "Figure 1. Maps of sampling locations. Dash circle indicating sadar upazila of Netrokona district and solid circle indicating Tarail upazila of Kishoreganj district. (Source: http://www.mapsofbangladesh.com/)",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/58/03/178-1597401409-Figure2.jpg",
"caption": "Figure 2. Results of propagation of present isolates of DPV in embryonated duck eggs. (a) normal embryo remained intact and CAM (arrow); (b) hemorrhagic thickened CAM and (c) petechial hemorrhages over the embryo’s body with no feather in DPV inoculated embryo; (d) Slide hemagglutination test mixing allantoic fluid from DPV infected embryo and 2% cRBC (chicken RBC) showed no hemagglutination.",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/58/03/178-1597401409-Figure3.jpg",
"caption": "Figure 3: Results of propagation of the present isolates of duck plague virus into duck embryo fibroblast cell culture. (a) Control: Growth pattern of duck embryo fibroblast cells (400X magnification), kept as control, showing confluent growth of star like fibroblast cells. (b) Infected: Observation CPE of duck embryo fibroblast cells by duck plague virus showing round, necrotic, degeneration of cells, clumping of cell and giant cell formation of fibroblast cells under green light (400X magnification)(arrow: clumping of cell; dash circle: giant cell; solid circle: round cell).",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/58/03/178-1597401409-Figure4.jpg",
"caption": "Figure 4. Results of pathogenicity test for the duck plague virus isolates. (a) Dead duckling after 6 days of post inoculation of DP virus; postmortem examination of ducklings showing (b) unclotted free blood in the body cavities, (c) petechial hemorrhage in the pale liver, (d) hemorrhagic annular band in intestine.",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/58/03/178-1597401409-Figure5.jpg",
"caption": "Figure 5. PCR amplification products of duck plague virus with DNA polymerase gene specific primer. Lane M: 100 bp ladder; Lane 1: positive control; Lane 2-6: positive samples of duck plague virus; Lane 7: negative control.",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/58/03/178-1597401409-Figure6.jpg",
"caption": "Figure 6. Comparative analysis of the nucleic acid sequenced data of the present isolates and Ahamed et al 2015 [10]. The nucleotide variation was found in the 374-379 position. The ‘solid box’ indicated the difference region and the ‘dash box’ indicated the nucleotide sequence which was not reported in the previous study from our laboratory (i.e., Ahamed et al., 2015, reported 378 bp out of 446 bp amplicon).",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/58/03/178-1597401409-Figure7.jpg",
"caption": "Figure 7. Phylogenetic tree showing relationship of DPV isolates. The tree was constructed through the neighbor joining method and nucleotide distances were measures through Jukes-Cantor method using CLC Sequence Viewer 8.0 software. Numbers on the nodes indicate bootstrap percentage values (calculated using 100 replicates). Red color represents this study isolates.",
"featured": false
}
],
"authors": [
{
"id": 480,
"affiliation": [
{
"affiliation": "Department of Microbiology and Hygiene, Bangladesh Agricultural University, Mymensingh-2202, Bangladesh"
}
],
"first_name": "Most. Tahmina",
"family_name": "Khan",
"email": null,
"author_order": 1,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 120
},
{
"id": 481,
"affiliation": [
{
"affiliation": "Department of Microbiology and Hygiene, Bangladesh Agricultural University, Mymensingh-2202, Bangladesh"
}
],
"first_name": "Md. Tahmid Rahman",
"family_name": "Pavel",
"email": null,
"author_order": 2,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 120
},
{
"id": 482,
"affiliation": [
{
"affiliation": "Department of Microbiology and Hygiene, Bangladesh Agricultural University, Mymensingh-2202, Bangladesh"
}
],
"first_name": "Afsana",
"family_name": "Keya",
"email": null,
"author_order": 3,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 120
},
{
"id": 483,
"affiliation": [
{
"affiliation": "Department of Microbiology and Hygiene, Bangladesh Agricultural University, Mymensingh-2202, Bangladesh"
}
],
"first_name": "Md. Ahosanul Haque",
"family_name": "Shahid",
"email": null,
"author_order": 4,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 120
},
{
"id": 484,
"affiliation": [
{
"affiliation": "Veterinary Surgeon, Department of Livestock Services, Peoples Republic of Bangladesh"
}
],
"first_name": "Tangila",
"family_name": "Ferdausi",
"email": null,
"author_order": 5,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 120
},
{
"id": 485,
"affiliation": [
{
"affiliation": "Department of Microbiology and Hygiene, Bangladesh Agricultural University, Mymensingh-2202, Bangladesh"
}
],
"first_name": "Mahbubul Pratik",
"family_name": "Siddique",
"email": null,
"author_order": 6,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 120
},
{
"id": 486,
"affiliation": [
{
"affiliation": "Department of Microbiology and Hygiene, Bangladesh Agricultural University, Mymensingh-2202, Bangladesh"
}
],
"first_name": "Muhammad Tofazzal",
"family_name": "Hossain",
"email": null,
"author_order": 7,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 120
},
{
"id": 487,
"affiliation": [
{
"affiliation": "Department of Microbiology and Hygiene, Bangladesh Agricultural University, Mymensingh-2202, Bangladesh"
}
],
"first_name": "KHM Nazmul Hussain",
"family_name": "Nazir",
"email": null,
"author_order": 8,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 120
},
{
"id": 488,
"affiliation": [
{
"affiliation": "Department of Microbiology and Hygiene, Bangladesh Agricultural University, Mymensingh-2202, Bangladesh"
}
],
"first_name": "Md. Tanvir",
"family_name": "Rahman",
"email": null,
"author_order": 9,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 120
},
{
"id": 489,
"affiliation": [
{
"affiliation": "Department of Microbiology and Hygiene, Bangladesh Agricultural University, Mymensingh-2202, Bangladesh"
}
],
"first_name": "Md. Bahanur",
"family_name": "Rahman",
"email": "bahanurr@bau.edu.bd",
"author_order": 10,
"ORCID": null,
"corresponding": true,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "Md. Bahanur Rahman, PhD; Professor, Department of Microbiology and Hygiene, Bangladesh Agricultural University, Mymensingh-2202, Bangladesh, Email: bahanurr@bau.edu.bd",
"article": 120
}
],
"views": 1049,
"downloads": 150,
"references": [
{
"id": 3880,
"serial_number": 1,
"pmc": null,
"reference": "Sarker AJ. Duck plague in Bangladesh: Isolation and identification of the etiological agent. Indian Vet J. 1982; 59: 669-679.",
"DOI": null,
"article": 120
},
{
"id": 3881,
"serial_number": 2,
"pmc": null,
"reference": "OIE. Manual of diagnosis tests and vaccines for terrestrial animals; OIE Terrestrial Manual, 2018. Chapter 2.3.7. Duck Virus Enteritis.",
"DOI": null,
"article": 120
},
{
"id": 3882,
"serial_number": 3,
"pmc": null,
"reference": "ICTV. Virus taxonomy. EC 47. London, UK: 2015. [Accessed on 16-10-2016]. Available from: http://www.ictvonline.org/virustaxonomy.asp.",
"DOI": null,
"article": 120
},
{
"id": 3883,
"serial_number": 4,
"pmc": null,
"reference": "Li Y, Huang B, Ma X, Wu J, Li F, Ai W et al. Molecular characterization of the genome of duck enteritis virus. Virology. 2009; 391(2): 151-161.",
"DOI": null,
"article": 120
},
{
"id": 3884,
"serial_number": 5,
"pmc": null,
"reference": "Cheng AC, Wang MS, Liu F, Song Y, Yuan GP, Han XY et al. Distribution and Excretion of Duck Plague Virus Attenuated Cha Strain in Vaccinated Ducklings [J]. Chinese J Vet. 2005; 3: 002.",
"DOI": null,
"article": 120
},
{
"id": 3885,
"serial_number": 6,
"pmc": null,
"reference": "Maclachlan NJ, Dubovi EJ, Fenner F (Eds). Fenner’s Veterinary Virology. 4th ed. Amsterdam: Elsevier Academic Press. 2011, pp: 1914-2010.",
"DOI": null,
"article": 120
},
{
"id": 3886,
"serial_number": 7,
"pmc": null,
"reference": "Hoque MA, Skerratt LF, Cook AJ, Khan SA, Grace D, Alam MR et al. Factors limiting the health of semi-scavenging ducks in Bangladesh. Trop Anim Health Prod. 2011; 43: 441-50.",
"DOI": null,
"article": 120
},
{
"id": 3887,
"serial_number": 8,
"pmc": null,
"reference": "Islam MR, Khan MAHNA. An immunocytochemical study on the sequential tissue distribution of duck plague virus. Avian Pathol. 1995; 24(1): 189-194.",
"DOI": null,
"article": 120
},
{
"id": 3888,
"serial_number": 9,
"pmc": null,
"reference": "Akter S, Islam MA, Hossain MT, Begum MIA, Amin MM, Sadekuzzaman M. Characterization and pathogenicity of Duck Plague virus isolated from natural outbreaks in ducks of Bangladesh. Bang J Vet Med. 2004; 2: 107-111.",
"DOI": null,
"article": 120
},
{
"id": 3889,
"serial_number": 10,
"pmc": null,
"reference": "Ahamed MM, Hossain MT, Rahman M, Nazir KHMNH, Khan MFR, Parvej MS, et al. Molecular characterization of Duck Plague virus isolated from Bangladesh. J Adv Vet Anim Res. 2015; 2(3): 296-303.",
"DOI": null,
"article": 120
},
{
"id": 3890,
"serial_number": 11,
"pmc": null,
"reference": "Hossain MT, Islam MA, Amin MM, Islam MA. Comparative efficacy of the conventional and experimentally developed duck plague vaccine. Int J Poult Sci. 2005; 4: 369-371.",
"DOI": null,
"article": 120
},
{
"id": 3891,
"serial_number": 12,
"pmc": null,
"reference": "El-Samadony HA, Tantawy LA, Salama E, Khedr AA. Molecular Characterization of Circulating Duck Viral Enteritis in Egypt during 2012-2013. Br J Poult Sci. 2013; 2: 38-44.",
"DOI": null,
"article": 120
},
{
"id": 3892,
"serial_number": 13,
"pmc": null,
"reference": "Gao X, Jia R, Wang M, Zhu D, Chen S, Lin M et al. Construction and identification of a cDNA library for use in the yeast two-hybrid system from duck embryonic fibroblast cells post-infected with duck enteritis virus. Mol Biol Rep. 2014; 41: 467-475.",
"DOI": null,
"article": 120
},
{
"id": 3893,
"serial_number": 14,
"pmc": null,
"reference": "Das SC, Chowdhury SD, Khatun MA, Nishibori M, Isobe N, Yoshimura Y. Poultry production profile and expected future projection in Bangladesh. World Poultry Sci J. 2008; 64: 99-116.",
"DOI": null,
"article": 120
},
{
"id": 3894,
"serial_number": 15,
"pmc": null,
"reference": "Wu Y, Cheng A, Wang M, Zhang S, Zhu D, Jia AB et al. Serologic detection of duck enteritis virus using an indirect ELISA based on recombinant UL55 protein. Avian Dis. 2011; 55: 626-632.",
"DOI": null,
"article": 120
},
{
"id": 3895,
"serial_number": 16,
"pmc": null,
"reference": "Hansen WR, Brown SE, Nashold SW, Knudson DL. Identification of duck plague virus by polymerase chain reaction. Avian Dis. 1999; 43(1): 106-115.",
"DOI": null,
"article": 120
},
{
"id": 3896,
"serial_number": 17,
"pmc": null,
"reference": "DLS. Annual report on livestock. 2018. Department of Livestock Services, Ministry of Fisheries and Livestock, Farmgate, Dhaka, Bangladesh",
"DOI": null,
"article": 120
},
{
"id": 3897,
"serial_number": 18,
"pmc": null,
"reference": "Hamid MA. Duck genetic resources, their improvement and conservation in Bangladesh: a review. SAARC J Agric. 2019; 17(2): 31-42.",
"DOI": null,
"article": 120
},
{
"id": 3898,
"serial_number": 19,
"pmc": null,
"reference": "CEGIS, Center for Environmental and Geographic Information Services. 2012. Master Plan of Haor Areas. Volume II, Main Report. Bangladesh Haor and Wetland Development Board, Ministry of Water Resources, Government of the Peoples’ Republic of Bangladesh. Pp: 109.",
"DOI": null,
"article": 120
},
{
"id": 3899,
"serial_number": 20,
"pmc": null,
"reference": "Cai MS, Cheng AC, Wang MS, Chen WP, Zhang X, Zheng SX et al. Characterization of the duck plague virus UL35 gene. Intervirology. 2010; 53(6), 408-416.",
"DOI": null,
"article": 120
},
{
"id": 3900,
"serial_number": 21,
"pmc": null,
"reference": "Hall TA. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. In Nucleic Acids Symposium Series 1999; (Vol. 41, No. 41, pp. 95-98). [London]: Information Retrieval Ltd., c1979-c2000.",
"DOI": null,
"article": 120
},
{
"id": 3901,
"serial_number": 22,
"pmc": null,
"reference": "Larkin MA, Blackshields G, Brown NP, Chenna R, McGettigan PA, McWilliam H et al. Clustal W and Clustal X version 2.0. Bioinformatics, 2007; 23(21), 2947-2948.",
"DOI": null,
"article": 120
},
{
"id": 3902,
"serial_number": 23,
"pmc": null,
"reference": "Hansen WR, Nashold SW, Docherty DE, Brown SE, Knudson DL. Diagnosis of duck plague in waterfowl by polymerase chain reaction. Avian Dis. 2000; 44: 266-274.",
"DOI": null,
"article": 120
},
{
"id": 3903,
"serial_number": 24,
"pmc": null,
"reference": "Soma SS, Nazir KHMNH, Rahman MT, Rahman MM, Ara MS, Sultana R et al. Isolation and Molecular Detection of duck plague virus for the development of vaccine seed. Asian Australas J Biosci Biotechnol. 2018; 3(1): 78-85.",
"DOI": null,
"article": 120
},
{
"id": 3904,
"serial_number": 25,
"pmc": null,
"reference": "Nguyen TKD, Ho VH, Nguyen TTH, Do VD, Tran DT, Nind L et al. Adaptation of duck plague virus to chicken embryo fibroblast cell culture for vaccine production. Proceedings, No. 117. In: Meers J, Spradbrow PB, DhinTu Tran, editors. Control of Newcastle disease and duck plague in village poultry. Canberra, Australia: Australian Center for International Agricultural Research; 2004. p. 53-56.",
"DOI": null,
"article": 120
},
{
"id": 3905,
"serial_number": 26,
"pmc": null,
"reference": "Aravind S, Kamble NM, Gaikwad SS, Shukla SK, Dey S, Mohan CM. Adaptation and growth kinetics study of an Indian isolate of virulent duck enteritis virus in Vero cells. Microb Pathog. 2015; 78: 14-19.",
"DOI": null,
"article": 120
},
{
"id": 3906,
"serial_number": 27,
"pmc": null,
"reference": "Zhang D, Lai M, Cheng A, Wang M, Wu Y, Yang Q et al. Molecular characterization of the duck enteritis virus US10 protein. Virol J. 2017; 14(1): 183.",
"DOI": null,
"article": 120
},
{
"id": 3907,
"serial_number": 28,
"pmc": null,
"reference": "Mandal PS, Mukhopadhayay SK, Pradhan S, Mondal S, Jana C, Patra NC et al. Development of nested polymerase chain reaction-based diagnosis of duck enteritis virus and detection of DNA polymerase gene from non-descriptive duck breeds of West Bengal, India. Vet World, 2017; 10(3): 336.",
"DOI": null,
"article": 120
}
]
},
{
"id": 117,
"slug": "178-1598378684-screening-of-antagonistic-potential-bacteria-from-rhizosphere-soil-against-phytopathogenic-fungi-related-to-selected-vegetable-crops",
"featured": false,
"slider": false,
"issue": "Vol4 Issue1",
"type": "original_article",
"manuscript_id": "178-1598378684",
"recieved": "2020-07-25",
"revised": null,
"accepted": "2020-10-01",
"published": "2020-10-08",
"pdf_file": "https://jabet.bsmiab.org/media/pdf_file/2023/14/178-1598378684.pdf",
"title": "Screening of antagonistic potential bacteria from rhizosphere soil against phytopathogenic fungi related to selected vegetable crops",
"abstract": "<p>Fungal phytopathogens cause serious losses of crop production worldwide, which causes serious losses of crop production. Bacteria play a role as the biocontrol agents for plant disease control. For this reason, the present study was conducted to determine the antagonistic potential of rhizosphere soil bacteria against selected phytopathogenic fungi. The screenings of potential antagonist isolated bacteria were applied by the dual culture technique with fungi <em>Fusarium oxysporum </em>and <em>Colletotrichum melongenae</em>. Molecular characterization was performed through 16S rDNA sequencing analysis. Sixteen (16) out of fifty (50) isolated bacteria showed different degrees of antagonism (25-67%) against both fungi <em>F. oxysporum </em>and <em>C. melongenae.</em> Among them, four (4) isolated bacteria (isolates A4, C1, C2, and E2) exhibited strong antagonism (more than 50% mycelial growth inhibition) against both fungi. The 16S rDNA sequences of the isolated bacteria A4, C1, C2 and E2 were 99-100% similar to <em>Providencia</em> sp. TT14, <em>Bacillus subtilis</em> 168, <em>Bacillus subtilis</em> RKP-2, and <em>Bacillus amyloliquefaciens</em> IBSDG-11, respectively. Based on the capability for the control of mycelial growth against both fungi, <em>B. subtilis</em> IUBTC2 was selected for optimization of growth characteristics and identification the bioactive metabolites which can enhance plant growth and disease control capacity of plants against the phytopathogenic fungi.</p>",
"journal_reference": "J Adv Biotechnol Exp Ther. 2021; 4(1): 35-43.",
"academic_editor": "Md Jamal Uddin, PhD; Ewha Womans University, Seoul, South Korea",
"cite_info": "Alam MK, Rahman MM, et al. Screening of antagonistic potential bacteria from rhizosphere soil against phytopathogenic fungi related to selected vegetable crops. J Adv Biotechnol Exp Ther. 2021; 4(1): 35-43.",
"keywords": [
"16S rDNA sequencing",
"Antagonistic bacteria",
"In vitro screening",
"Phytopathogenic fungi",
"Vegetable crops"
],
"DOI": "10.5455/jabet.2021.d104",
"sections": [
{
"section_number": 1,
"section_title": "INTRODUCTION",
"body": "<p>Major crop production losses are caused worldwide by pathogenic microorganisms which is comparable to approximately $220 billion lost per year [<a href=\"#r-1\">1</a>]. Worldwide, the crop production damage was also caused by different fungal phytopathogenic species as <em>Fusarium</em>, <em>Rhizoctonia</em> or <em>Colletotrichum</em> such [<a href=\"#r-2\">2-5</a>]. <em>Fusarium</em> wilt of cucumber diseases was caused by <em>F. oxysporum, </em>which is a very common disease in cucumber worldwide and causes serious losses every year [<a href=\"#r-6\">6-8</a>]. Until now, disease control mechanism mostly depends on the practice of artificial fungicides. This approach is very challenging, because fungal stains may become resistant to fungicides due to the accumulation of these artificial fungicide’s in the ecosystem [<a href=\"#r-9\">9</a>, <a href=\"#r-3\">3</a>]. Artificial fungicides are expensive. So, these are not sufficient for the disease control strategies [<a href=\"#r-10\">10</a>].<br />\r\nIn addition, public anxiety related to chemical pesticides has raised key attention to pursue additional control approaches [<a href=\"#r-11\">11</a>]. Interactions are prevalent in nature between plant pathogens and antagonistic microorganisms. Similarly, plant pathogenic fungi and antagonistic microorganism interactions can employ to control or reduce diseases as biocontrol/biological agents which can suppress the plant pathogens [<a href=\"#r-12\">12</a>]. It is one of the paths to reduce the usage of artificial fungicides in agriculture sector, where plant protection from diseases can be done by using microorganisms [<a href=\"#r-13\">13</a>]. The biocontrol or biological control mechanisms are the secretion of both small molecules and enzymes, which are extremely active against pathogens [<a href=\"#r-14\">14</a>, <a href=\"#r-1\">1</a>, <a href=\"#r-3\">3</a>].<br />\r\nAntagonism is the phenomenon in which one microorganism inhibits the other interacting partner to ensure its own survival. The stalling products of microbes are preferably called antibiotic substances or metabolites. It was reported that antagonistic bacteria act as a biocontrol agent due to their ability of the different modes of action [<a href=\"#r-12\">12</a>]. The antagonistic effect of microorganism is the result of interaction among the microbial populations. Several bacterial species have been tested as biocontrol/biological agents [<a href=\"#r-15\">15</a>]. The bacterial antagonisms mechanisms are involved in competition for space or nutrients and antibiosis for enrichment of plant and root growth, and for introduction of plant inactivation and/or resistance of the pathogen’s enzymes [<a href=\"#r-16\">16</a>].<br />\r\nAntibiosis is the utmost significant machinery for the plant disease control. Cucumber (<em>Cucumis sativus</em> L.) is one of the key significant economical crops, which is grown either in the open field or greenhouses [<a href=\"#r-17\">17, 18</a>]. <em>Fusarium</em> wilt of cucumber, caused by <em>F. oxysporum</em> <em>f. sp. cucumerinum</em>, is a very common disease of cucumber and oil palm in the world and causes serious losses every year [<a href=\"#r-19\">19</a>, <a href=\"#r-6\">6</a>]. Eggplant (<em>Solanum melongena</em>) is a tropical, herbaceous, perennial plant, in the family Solanaceae, which is grown for its edible fruit [<a href=\"#r-20\">20</a>]. The marketable eggplant production is negotiated due to affect by the insect which are generally recognized such as eggplant shoot and fruit borer (<em>Leucinodes orbonalis</em>) as well as various phytopathogenic fungi namely, <em>Fusarium </em>spp., <em>C. melongenae</em>, <em>Pythium</em> spp., <em>Leveillu lataurica,</em> <em>Rhizoctonia</em> spp., and <em>Verticillium</em> spp<em>. </em>[<a href=\"#r-17\">17</a>]. Until now, disease control mainly deepens on the usage of synthetic fungicides. For this reason, the objective of this study was to screen the antagonistic potential of bacteria from rhizosphere soil against phytopathogenic fungi in selected vegetable crops in Bangladesh. At first, we isolated bacteria from rhizosphere soil at different sources and screened the antagonistic potential bacteria against phytopathogenic fungi. We also further investigated as morphological, biochemical and molecular characterization to determine the biocontrol/biological agents for the phytopathogenic fungus control.</p>"
},
{
"section_number": 2,
"section_title": "MATERIALS AND METHODS",
"body": "<p><strong>Fungal pathogens</strong><br />\r\nTwo fungal pathogens, namely, <em>F. oxysporum cucumerinum</em> and <em>C. melongenae</em> were used in this study, which were obtained from the Laboratory of Microbiology, Islamic University, Kushtia-7003, Bangladesh.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Isolation of soil </strong><strong>bacteria</strong><br />\r\nRhizosphere soil was collected from 5 different agricultural and non-agricultural field crops of Shantidanga, Kushtia. The soil samples were brought to the laboratory in sterile polyethylene bags and stored at 4ºC for the isolation of microorganisms. Soil bacteria were isolated by serial dilution method from soil samples. In brief, it was suspended 5 g of soil in sterile distilled water and shaking at 120 rpm for 10 min in a rotary shaker. Then it was diluted with distilled water (1:9 ratio) up to10<sup>7 </sup>fold. 100 μl aliquots from 10<sup>4 </sup>to 10<sup>7</sup> dilutions was spread in the tryptone soya agar (TSA) media and mildly spread by a sterile glass spreader. Then it was incubated at 35ºC for 3 days, after it was sub-cultured onto the same medium plates based on morphological distinct colony and isolated single colonies from these plates.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Screening of antagonistic activities</strong><br />\r\nAntagonistic activities were screened from the isolated bacterial strains through <em>In vitro.</em> Mycelial growth inhibition of <em>F. oxysporum </em>and <em>C. melongenae</em> was achieved through the dual culture method in potato dextrose agar (PDA) media. PDA media was organized and transferred in petri dishes (20 ml). Agar plug (5 mm) of vigorously rising culture of each fungus was positioned into the center of individual plate. Individual isolate was streaked 3 cm away and 1 cm away from the “edge of the” petri dish. Plates without bacterial inoculation was used a control. Then the plate was enclosed with parafilm and incubated at a temperature (27ºC) for 5 days till the fungal mycelia touched the control plate edge. All tests were done in triplicate for each isolate. Then circular mycelial development, inhibition percentage (%) was measured by the following formula [<a href=\"#r-21\">21</a>]:<br />\r\nRadial inhibition (RI%)=(A1-A2)/A1x100<br />\r\nWhere, A1 = radial growth of fungal mycelia without bacterial isolate, A2 = radial growth of fungal mycelia with bacterial isolate.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Morphological characterization and biochemical test of isolated bacteria</strong><br />\r\nThe morphological characterizations and physiological and biochemical tests were done on the selected isolates. In brief, morphological characterizations such as size, shape, motility and spore forming were observed. The biochemical tests such as Gram staining, catalase test, KOH test, lactose fermentation, sulfur reduction, and urease test of the isolated bacteria were done as described earlier [<a href=\"#r-22\">22</a>, <a href=\"#r-23\">23</a>]. Determination of degree of growth of isolated bacteria, different types of media were used such as tryptone soya agar (TSA), lactose broth agar (LBA), yeast extract agar (YEA), potato dextrose agar (PDA), and King’s medium B agar (KBA).</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Molecular characterization of the isolate</strong><strong>d bacteria</strong><br />\r\nGenomic DNA was extracted from the selected isolated bacteria according to the protocol designated by He [<a href=\"#r-24\">24</a>] (phenol: chloroform: iso-amyl alcohol procedure). The sequence of 16S rDNA was amplified through polymerase chain reaction (PCR). Thermal cycles and other conditions were applied according to Rahman et al [<a href=\"#r-25\">25, 26</a>]. The universal primer set of 63F (5-CAGGCCTAACACATGCAAGTC-3) [<a href=\"#r-25\">25, 26, 27</a>] and 1389R (5-ACGGGCGGTGTGTACAAG-3) [<a href=\"#r-25\">25, 26</a>, <a href=\"#r-28\">28</a>] was used for the amplification of 16S rDNA. Amplified PCR products were purified by purification kits (TaKaRa, Hot-Start Version). Then the purified PCR products were sequenced by DNA sequencer (Invent Technology, Dhaka, Bangladesh). The obtained sequences were analyzed through Chromas software. The similarity was observed with known sequences present in the NCBI GenBank database (<a href=\"http://blast.ncbi.nlm.nih.gov/Blast.cgi\">http://blast.ncbi.nlm.nih.gov/Blast.cgi</a>) through the BLAST program. Multiple sequence alignment (MSA) of 16S rDNA sequences of the isolated bacteria was performed by using the Clustal Omega (<a href=\"https://www.ebi.ac.uk/Tools/msa/clustalo/\">https://www.ebi.ac.uk/Tools/msa/clustalo/</a>). The aligned sequences were then exposed to a Neighbor-Joining phylogenetic tree constructed by Clustal Omega. Then 16S r DNA sequences were deposited into the NCBI data bank. The sequences were submitted in the NCBI gene bank and accession numbers as MG237885 to MG237888 for <em>Providencia</em> sp. IUBTA4, <em>B. subtilis</em> IUBTC1, <em>B. subtilis</em> IUBTC2, and <em>B. amyloliquefaciens</em> IUBTE2, respectively.</p>"
},
{
"section_number": 3,
"section_title": "RESULTS",
"body": "<p><strong>Screening of antagonistic activities from </strong><strong>isolated bacteria through an in vitro assay</strong><br />\r\nScreening of antagonistic activity of each isolated bacterium was accomplished through an<em> in vitro </em>assay as shown in <a href=\"#figure1\">Figure 1</a>. Fifty (50) bacterial isolates were obtained from five (5) different soil samples. Some isolates were antagonists against only one fungus. Among them, sixteen (16) isolates showed different degrees of antagonism (25-67%) against both fungi, <em>F. oxysporum </em>and <em>C. melongenae</em>. Four (4) isolates A2 (58% and 57%), C1 (50% and 63%), C2 (67% and 64%), and E2 (62% and 50%) showed a higher efficacy in inhibiting the radial growth fungal mycelia. The strong antagonism (more than 50% mycelial growth inhibition) was exhibited by the isolates A4, C1, C2, and E2 against both fungi (<a href=\"#figure2\">Figure 2</a>)<em>.</em></p>\r\n\r\n<div id=\"figure1\">\r\n<figure class=\"image\"><img alt=\"\" height=\"220\" src=\"/media/article_images/2024/41/03/178-1598378684-Figure1.jpg\" width=\"485\" />\r\n<figcaption><strong>Figure 1. </strong> Antagonistic activity of isolated bacteria against <em>F. oxysporum</em> and <em>C. melongenae</em> in dual culture in PDA after 4 days of culture at 27ºC. (a-d and f-i). Fungal growth was inhibited towards the direction of isolated bacteria and (e and j) control plates inoculated with only the fungal isolate.</figcaption>\r\n</figure>\r\n</div>\r\n\r\n<div id=\"figure2\">\r\n<figure class=\"image\"><img alt=\"\" height=\"412\" src=\"/media/article_images/2024/41/03/178-1598378684-Figure2.jpg\" width=\"481\" />\r\n<figcaption><strong>Figure 2. </strong> Antagonistic activities of potential isolated bacteria against <em>F. oxysporum </em>and <em>C. melongenae. </em>Selected bacteria, A4-<em> Providencia </em>spp. TT14, C1-<em> B. subtilis </em>168<em>, </em>C2-<em> B. subtilis</em> RKP-2, and E2-<em>B. amyloliquefaciens</em> IBSDG-11.</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Morphological characterization of isolated bacteria</strong><br />\r\nMorphological characterizations of isolated bacteria are shown in <a href=\"#Table-1\">Table 1</a>. Morphological studies showed that the isolated bacteria A4 was straight rods, non-spore former, and motile. The isolated bacteria C1, C2, and E2 were rods shape, spore former, and motile. The degree of growth characteristics of the isolated bacteria A4, C1, C2 and E2 was observed using different types of media as shown in <a href=\"#Table-2\">Table 2</a> and the growth patterns were recorded as shown <a href=\"#figure3\">Figure 3</a>. Four (4) isolated bacteria A4, C1, C2 and E2 showed the excellence growth and good growth on TSA and KBA media, respectively as compared with other media. The growth patterns revealed that there was a distinct difference among the isolated bacteria.</p>\r\n\r\n<div id=\"figure3\">\r\n<figure class=\"image\"><img alt=\"\" height=\"399\" src=\"/media/article_images/2024/41/03/178-1598378684-Figure3.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 3.</strong> Growth patterns of the selected isolated bacteria. Selected bacteria, A4- <em>Providencia</em> spp. TT14, C1- <em>B. subtilis </em>168, C2- <em>B. subtilis</em> RKP-2, and E2-<em>B. amyloliquefaciens</em> IBSDG-11in different culture media (a) Tryptone soya agar (TAS), (b) lactose broth agar (LBA), (c) yeast extract agar (YEA), (d) potato dextrose agar (PDA) and (e) King’s medium B agar (KBA) after 66 h of incubation at 37ºC</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<div id=\"Table-1\">\r\n<p><a href=\"https://jabet.bsmiab.org/table/178-1598378684-table1/\">Table-1</a><strong>Table 1. </strong>Morphological characteristics and the biochemical properties of the selected isolated bacteria.</p>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<div id=\"Table-2\">\r\n<p><a href=\"https://jabet.bsmiab.org/table/178-1598378684-table2/\">Table-2</a><strong>Table 2. </strong>Degree of growth of the selected isolated bacteria in different culture media.</p>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Biochemical test of isolated bacteria</strong><br />\r\nThe results of biochemical test of isolated bacteria were shown in <a href=\"#Table-2\">Table 2</a>. The isolated bacteria C1, C2, and E2 were positive for the Gram staining and KOH test, respectively. Isolated bacteria A4 was negative against gram reaction and KOH test, respectively. Isolated bacteria A4, C1, C2, and E2 were positive in the catalase test and urease test. On the other hand, all of the four isolated bacteria showed negative for lactose fermentation and sulphur reduction (<a href=\"#Table-2\">Table 2</a>).</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Molecular identification of selected bacterial isolated</strong><br />\r\nMolecular identification was applied by 16S rDNA sequence analysis as shown in <a href=\"#Table-3\">Table 3</a>. The 16S rDNA sequences were received and analyzed in the Chromas software (version 2.6.4) showed good quality of the sequences. The similarity of the sequence of the isolates was exposed (100-99% similarity) through BLAST exploration and there was a close relationship presented between the isolated strains and known sequence of the gene bank NCBI. Isolated bacteria A4, C1, C2 and E2 were most closely related to <em>Providencia</em> sp. BAB-5310<em>, Bacillus</em> sp. E157, <em>B. subtilis</em> subsp. <em>inaquosorum</em> strain PF9 and <em>B. amyloliquefaciens</em> strain BG2. The sequences were submitted in the NCBI Gene Bank and accession numbers A4 (MG237885), C1 (MG237886), C2 (MG237887) and E2 (MG237888) were received for the isolated bacteria. Finally, the four isolates were identified as <em>Providencia </em>sp. IUBTA4, <em>B. subtilis </em>IUBTC1, <em>B. subtilis </em>IUBTC2 and <em>B. amyloliquefaciens </em>IUBTE2. Neighbor-joining phylogenetic trees were constructed of isolated bacteria as shown in <a href=\"#figure4\">Figure 4</a>. Phylogenetic analysis indicated that isolated bacteria were represented in a distinct class, demonstrating that they are newly isolated bacteria as compared to NCBI Data bank.</p>\r\n\r\n<div id=\"figure4\">\r\n<figure class=\"image\"><img alt=\"\" height=\"685\" src=\"/media/article_images/2024/41/03/178-1598378684-Figure4.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 4. </strong>Neighbor-joining phylogenetic trees showing relationships of isolated bacteria a) A4- <em>Providencia </em>sp. TT14, b) C1- <em>B. subtilis</em> 168, c) C2<em>– B. subtilis</em> RKP-2, and d) E2-<em>B.</em> <em>amyloliquefaciens</em> IBSDG-11 with closely related strains based on their 16S rDNA sequences</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<div id=\"Table-3\">\r\n<p><a href=\"https://jabet.bsmiab.org/table/178-1598378684-table3/\">Table-3</a><strong>Table 3. </strong>Closely related species of the four selected isolated bacteria based on the similarity of the partial 16S rDNA sequences in BLASTN.</p>\r\n\r\n<p> </p>\r\n</div>"
},
{
"section_number": 4,
"section_title": "DISCUSSION",
"body": "<p>The use of valuable microorganisms considers the greatest auspicious approaches for the harmless crop management performs [<a href=\"#r-29\">29</a>]. In this respect, we tried to isolate and identify antagonistic bacteria with robust antifungal activities against <em>F. oxysporum </em>and <em>C. melongenae</em>. In our investigation, we found that among fifty (50) isolated bacteria, sixteen (16) isolated bacteria showed different degrees of antagonism (25-67%) against both fungi in dual culture. Four isolates with an inhibition rate above 50% were selected for further investigations (<a href=\"#figure2\">Figure 2</a>). <em>In vitro</em> dual culture examination was widely applied to the preliminary screening of biocontrol/biological agents [<a href=\"#r-21\">21</a>]. Effects of antagonistic are frequently established by the development of inhibition zones between fungal and isolated bacteria [<a href=\"#r-21\">21</a>] or by calculating the radial mycelial growth inhibition in percentage towards the isolated bacteria [<a href=\"#r-30\">30</a>]. The selected isolates were identified based on morphological, biochemical and molecular techniques.<br />\r\nMolecular characterization was established and approved almost 2 decades years before to identify isolated bacteria species. This identification is commonly based on some sole parts of their subunit of 30S ribosomal RNA, which is called 16S rDNA [<a href=\"#r-31\">31</a>]. So, 16S rDNA- sequence based techniques were extensively applied to characterize the bacterial community structure in soils [<a href=\"#r-32\">32</a>, <a href=\"#r-25\">25</a>, <a href=\"#r-26\">26</a>, <a href=\"#r-33\">33</a>]. The 16S rDNA sequences of isolateA4, C1, C2, and E2 revealed that they were closely matched with <em>Providencia </em>sp, <em>B. subtilis</em> 168, <em>B. subtilis</em> RKP-2, and<em> B. amyloliquefaciens, </em>respectively (<a href=\"#Table-3\">Table 3</a>).</p>\r\n\r\n<p>The <em>B. subtilis</em> are Gram positive, rod shaped, motile soil bacteria. Both <em>B. subtilis</em> strains showed the antagonistic activities in the dual culture. <em>B. subtilis </em>IUBTC1 showed the inhibition rates 50% and 63% against fungi <em>F. oxysporum </em>and <em>C. melongenae </em>respectively, whereas <em>B. subtilis </em>IUBTC2 showed the inhibition rates 67% and 64% against both fungi respectively due to produce more enzyme substances (<a href=\"#figure2\">Figure 2</a>). Similarly, Burhan et al. [<a href=\"#r-34\">34</a>] demonstrated that different species of <em>Bacillus</em>, most notably, <em>B. subtilis</em> produced approximately 60% of commercially available enzymes. In previous studies, <em>Bacillus </em>spp. exhibited very broad spectra of action with an efficient antagonistic activity against <em>F. oxysporum </em>[<a href=\"#r-35\">35-37</a>]. Several species of <em>Bacillus</em> have been recognized as plant-growth promoting bacteria (PGPB) and/or biocontrol agents (BCA) [<a href=\"#r-37\">37</a>]. It was also reported that the genus <em>Bacillus</em> has become a dependable choice to discovery out novel and promising bacteria for the making of amylase and other extracellular enzymes [<a href=\"#r-33\">33</a>]. According to the NCBI database, isolate E2 was identified as <em>B. amyloliquefaciens </em>IUBTE2. It exhibited inhibition rates 62% and 50% against <em>F. oxysporum </em>and <em>C. melongenae, </em>respectively (<a href=\"#figure2\">Figure 2</a>). In previous, it was reported that the non-pathogenic bacilli, <em>B. amyloliquifaciens</em> showed an inhibition rate of 46% against <em>F. oxysporum </em>[<a href=\"#r-38\">38</a>]. <em>Providencia </em>spp. are as one of the most common multi-drug resistant bacteria [<a href=\"#r-39\">39</a>]. Similarly, Godebo et al [<a href=\"#r-40\">40</a>] reported that 75% of <em>Providencia </em>isolates were multi-drug resistant. In this study, isolated bacteria A4 was identified as <em>Providencia </em>sp. IUBTA4 that showed 58% and 57% mycelial growth inhibition against <em>F. oxysporum </em>and <em>C. melongenae, </em>respectively, due to biocontrol potential elicited defense enzymes (<a href=\"#figure2\">Figure 2</a>). Similarly, Rana et al. [<a href=\"#r-41\">41</a>] reported that <em>Providencia </em>spp. act as PGPR with biocontrol potential elicited defense enzymes in wheat.</p>"
},
{
"section_number": 5,
"section_title": "CONCLUSION",
"body": "<p>The main ecological tasks facing microbiologists and plant pathologists are to produce the environment-friendly substitutions instead of chemical pesticides for the fighting of crop infections. Sixteen (16) isolates showed different degrees of antagonism (25-67%) against both fungi <em>F. oxysporum</em> and <em>C. melongenae</em>. Four (4) isolated bacteria, <em>Providencia</em> sp. TT14, <em>B. subtilis</em> 168, <em>B. subtilis</em> RKP-2, and <em>B. amyloliquefaciens</em> IBSDG-11, respectively were exhibited strong antagonism (more than 50% mycelial growth inhibition). <em>B. subtilis</em> IUBTC2 showed the highest capability for the control of mycelial growth against phytopathogenic fungi. Finally, it may conclude that newly isolated bacteria have a potential disease control capacity of the plant against the pathogenic fungi which can be enhanced plant growths and seed emergences as well as act as biocontrol agents for better green environmental management (<a href=\"#figure5\">Figure 5</a>).</p>\r\n\r\n<div id=\"figure5\">\r\n<figure class=\"image\"><img alt=\"\" height=\"432\" src=\"/media/article_images/2024/41/03/178-1598378684-Figure5.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 5.</strong> Schematic diagram of summary.</figcaption>\r\n</figure>\r\n</div>"
},
{
"section_number": 6,
"section_title": "ACKNOWLEDGMENTS",
"body": "<p>The authors grateful to Department of Biotechnology and Genetic Engineering, Islamic University, Kushtia-7003, Bangladesh for assistance of research activities.</p>"
},
{
"section_number": 7,
"section_title": "AUTHOR CONTRIBUTIONS",
"body": "<p>MKA, MMR, and MRI comprehended and planned the study; MKA, and AHMJ carried out the analysis; MKA, and MMR wrote the manuscript; MMR prepared the graphs and illustrations; AHMJ and MRI contributed to the critical revision of the manuscript; MMR and MRI supervised the whole work; and all authors approved the final manuscript.</p>"
},
{
"section_number": 8,
"section_title": "CONFLICTS OF INTEREST",
"body": "<p>There is no conflict of interest among the authors.</p>"
}
],
"figures": [
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/41/03/178-1598378684-Figure1.jpg",
"caption": "Figure 1. Antagonistic activity of isolated bacteria against F. oxysporum and C. melongenae in dual culture in PDA after 4 days of culture at 27ºC. (a-d and f-i). Fungal growth was inhibited towards the direction of isolated bacteria and (e and j) control plates inoculated with only the fungal isolate.",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/41/03/178-1598378684-Figure2.jpg",
"caption": "Figure 2. Antagonistic activities of potential isolated bacteria against F. oxysporum and C. melongenae. Selected bacteria, A4- Providencia spp. TT14, C1- B. subtilis 168, C2- B. subtilis RKP-2, and E2-B. amyloliquefaciens IBSDG-11.",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/41/03/178-1598378684-Figure3.jpg",
"caption": "Figure 3. Growth patterns of the selected isolated bacteria. Selected bacteria, A4- Providencia spp. TT14, C1- B. subtilis 168, C2- B. subtilis RKP-2, and E2-B. amyloliquefaciens IBSDG-11in different culture media (a) Tryptone soya agar (TAS), (b) lactose broth agar (LBA), (c) yeast extract agar (YEA), (d) potato dextrose agar (PDA) and (e) King’s medium B agar (KBA) after 66 h of incubation at 37ºC",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/41/03/178-1598378684-Figure4.jpg",
"caption": "Figure 4. Neighbor-joining phylogenetic trees showing relationships of isolated bacteria a) A4- Providencia sp. TT14, b) C1- B. subtilis 168, c) C2- B. subtilis RKP-2, and d) E2-B. amyloliquefaciens IBSDG-11 with closely related strains based on their 16S rDNA sequences.",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/41/03/178-1598378684-Figure5.jpg",
"caption": "Figure 5. Schematic diagram of summary.",
"featured": false
}
],
"authors": [
{
"id": 472,
"affiliation": [
{
"affiliation": "Department of Biotechnology and Genetic Engineering, Islamic University, Kushtia-7003, Bangladesh."
}
],
"first_name": "Md. Khasrul Alam",
"family_name": "Alam",
"email": null,
"author_order": 1,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 117
},
{
"id": 473,
"affiliation": [
{
"affiliation": "Department of Biotechnology and Genetic Engineering, Islamic University, Kushtia-7003, Bangladesh."
}
],
"first_name": "M Mizanur",
"family_name": "Rahman",
"email": "rahmanmm@btge.iu.ac.bd",
"author_order": 2,
"ORCID": "https://orcid.org/0000-0003-4225-9451",
"corresponding": true,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "Prof. Dr. Md. Mizanur Rahman, Department of Biotechnology and Genetic Engineering, Islamic University, Kushtia-7003, Bangladesh. E-mail: rahmanmm@btge.iu.ac.bd",
"article": 117
},
{
"id": 474,
"affiliation": [
{
"affiliation": "Department of Biotechnology and Genetic Engineering, Islamic University, Kushtia-7003, Bangladesh."
}
],
"first_name": "Abu Hena Mostafa",
"family_name": "Jamal",
"email": null,
"author_order": 3,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 117
},
{
"id": 475,
"affiliation": [
{
"affiliation": "Department of Biotechnology and Genetic Engineering, Islamic University, Kushtia-7003, Bangladesh."
}
],
"first_name": "Md. Rezuanul",
"family_name": "Islam",
"email": null,
"author_order": 4,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 117
}
],
"views": 1566,
"downloads": 282,
"references": [
{
"id": 3773,
"serial_number": 1,
"pmc": null,
"reference": "Kandel SL, Firrincieli A, Joubert PM, Okubara PA, Leston ND, McGeorge KM, et. al, An in vitro study of biocontrol and plant growth promotion potential of Salicaceae endophytes. Frontiers in Microbiology. 2017; 8:386.",
"DOI": null,
"article": 117
},
{
"id": 3774,
"serial_number": 2,
"pmc": null,
"reference": "Imran QM, Yun B. Pathogen-induced Defense Strategies in Plants. Journal of Crop Science and Biotechnology.2020; 23:97–105,",
"DOI": null,
"article": 117
},
{
"id": 3775,
"serial_number": 3,
"pmc": null,
"reference": "Mehnaz S, Saleem RSZ, Yameen B, Pianet I, Schnakenburg G, Pietraszkiewicz H, Harald G. Lahorenoic acids A-C, ortho-dialkyl-substituted aromatic acids from the biocontrol strain Pseudomonas aurantiaca PB-St2. Journal of Natural Products 2013;76(2):135–141.",
"DOI": null,
"article": 117
},
{
"id": 3776,
"serial_number": 4,
"pmc": null,
"reference": "Winterab M, Samuelsa PL, Donga Y, Dill-Mackya R. Trichothecene production is detrimental to early root colonization by Fusarium culmorum and F. graminearum in fusarium crown and root rot of wheat. Plant Pathology. 2019; 68:185–19.",
"DOI": null,
"article": 117
},
{
"id": 3777,
"serial_number": 5,
"pmc": null,
"reference": "Patel S, Rajput K, Saraf M. Elicitation of plant defense enzymes against Fusarium oxysporum f. sp. lycopersici in tomato plant using a novel rhizobacteria Providencia rettgeri MSS2. Biocatalysis and Agricultural Biotechnology. 2017; 12:308313.",
"DOI": null,
"article": 117
},
{
"id": 3778,
"serial_number": 6,
"pmc": null,
"reference": "Kazan K, Gardiner DM. Transcriptomics of cereal-Fusarium graminearum interactions: what we have learned so far. Mol. Plant Pathology. 2018; 19(3):764–778.",
"DOI": null,
"article": 117
},
{
"id": 3779,
"serial_number": 7,
"pmc": null,
"reference": "Das MM, Haridas M, Sabu A. Biological control of black pepper and ginger pathogens, Fusarium oxysporum, Rhizoctonia solani and Phytophthora capsici, using Trichoderma spp. Biocatalysis and Agricultural Biotechnology. 2019; 17:177–183.",
"DOI": null,
"article": 117
},
{
"id": 3780,
"serial_number": 8,
"pmc": null,
"reference": "Palacio-Barrera AM, Areiza D, Zapata P, Atehortúa L, Correa C, Peñuela-Vásquez M. Induction of pigment production through media composition, abiotic and biotic factors in two filamentous fungi. Biotechnology Reports. 2019; 21:e00308.",
"DOI": null,
"article": 117
},
{
"id": 3781,
"serial_number": 9,
"pmc": null,
"reference": "Hassan MN, Afghan S, Hafeez FY. Biological control of red rot in sugarcane by native pyoluteorin-producing Pseudomonas putida strain NH-50 under field conditions and its potential modes of action. Pest Management Science. 2011; 67(9):1147–1154.",
"DOI": null,
"article": 117
},
{
"id": 3782,
"serial_number": 10,
"pmc": null,
"reference": "Kumar A, Verma H, Singh VK, Singh PP, Singh SK, Ansari WA, Pandey KD. Role of Pseudomonas sp. in sustainable agriculture and disease management. In V. S. Meena, P. K. Mishra, J. K. Bisht, & A. Pattanayak (Eds.), Agriculturally Important Microbes for Sustainable Agriculture Singapore: Springer Singapore (2017). doi.10.1007/978-981-10-5343-6_7",
"DOI": null,
"article": 117
},
{
"id": 3783,
"serial_number": 11,
"pmc": null,
"reference": "Nicolopoulou-Stamati P, Maipas S, Kotampasi C, Stamatis P, Hens L. Chemical pesticides and human health: the urgent need for a new concept in agriculture. Frontiers in Public Health. 2016; 4 :148",
"DOI": null,
"article": 117
},
{
"id": 3784,
"serial_number": 12,
"pmc": null,
"reference": "Köhl J, Kolnaar R, Ravensberg WJ. Mode of action of microbial biological control agents against plant diseases: relevance beyond efficacy. Frontiers in Plant Science. 2019; 10: 845.",
"DOI": null,
"article": 117
},
{
"id": 3785,
"serial_number": 13,
"pmc": null,
"reference": "van Lenteren JC, Bolckmans K, Köhl J, Ravensberg WJ, Urbaneja A. Biological control using invertebrates and microorganisms: plenty of new opportunities. BioControl. 2018; 63: 39–59.",
"DOI": null,
"article": 117
},
{
"id": 3786,
"serial_number": 14,
"pmc": null,
"reference": "Eilenberg J, Hajek A, Lomer C. Suggestions for unifying the terminology in biological control. BioControl. 2001; 46:387–400.",
"DOI": null,
"article": 117
},
{
"id": 3787,
"serial_number": 15,
"pmc": null,
"reference": "Moita C, Feio SS, Nunes L, Curto MJM, Roseiro JC. Optimization of physical factors on the production of active metabolites by Bacillus subtilis 355 against wood surface contaminant fungi. International Biodeterioration & Biodegradation. 2005; 55(4): 261–269.",
"DOI": null,
"article": 117
},
{
"id": 3788,
"serial_number": 16,
"pmc": null,
"reference": "Harman GE. Myths and Dogmas of biocontrol changes in perceptions derived from research on Trichoderma harzinum T-22. Plant Disease. 2000; 84(4):377–393.",
"DOI": null,
"article": 117
},
{
"id": 3789,
"serial_number": 17,
"pmc": null,
"reference": "Islam MA, Nain Z, Alam MK. et al. In vitro study of biocontrol potential of rhizospheric Pseudomonas aeruginosa against Fusarium oxysporum f. sp. cucumerinum. Egyptian Journal of Biological Pest Control. 2018; 28:90.",
"DOI": null,
"article": 117
},
{
"id": 3790,
"serial_number": 18,
"pmc": null,
"reference": "Saravanakumar K, Yu C, Dou K, Wang M, Li Y, Chen J. Synergistic effect of Trichoderma-derived antifungal metabolites and cell wall degrading enzymes on enhanced biocontrol of Fusarium oxysporum f. sp. cucumerinum. Biological Control. 2016; 94:37–46.",
"DOI": null,
"article": 117
},
{
"id": 3791,
"serial_number": 19,
"pmc": null,
"reference": "Adusei-Fosu K, Dickinson M, Yankey EN. AFLP as a fingerprinting tool for characterising isolates of Fusarium oxysporum f. sp. elaeidis causal organism for fusarium wilt disease of oil palm in Ghana. Journal of Plant Diseases and Protection. 2019; 126:575–584.",
"DOI": null,
"article": 117
},
{
"id": 3792,
"serial_number": 20,
"pmc": null,
"reference": "Parvaiz M, Hussain K, Rafique Y, Haider A, Nasreen S, Kousar S. In vitro Techniques for Propagation of Brinjal (Solanum melongena L.). World Applied Sciences Journal. 2013; 27(8) :1071–1078.",
"DOI": null,
"article": 117
},
{
"id": 3793,
"serial_number": 21,
"pmc": null,
"reference": "Ji SH, Gururani MA, Chun SC. Isolation and characterization of plant growth promoting endophytic diazotrophic bacteria from Korean rice cultivars. Microbiological Research. 2014; 169(1):83–98.",
"DOI": null,
"article": 117
},
{
"id": 3794,
"serial_number": 22,
"pmc": null,
"reference": "Rahman MM, Rahman MM, Akhter S, Jamal MA, Pandeya DR, Haque MA, Alam MF, Rahman A. Control of coliform bacteria detected from diarrhea associated patients by extracts of Moringa oleifera. Nepal Medical College Journal. 2010; 12:12–19.",
"DOI": null,
"article": 117
},
{
"id": 3795,
"serial_number": 23,
"pmc": null,
"reference": "Chomvarin C, Chantarasuk Y, Mairiang P. et al. Sensitivity and specificity of an in-house rapid urease test for detecting Helicobacter pylori infection on gastric biopsy. The Southeast Asian Journal of Tropical Medicine and Public Health. 2006; 37(2):312–9.",
"DOI": null,
"article": 117
},
{
"id": 3796,
"serial_number": 24,
"pmc": null,
"reference": "He F. E. coli Genomic DNA Extraction. Bio-protocol. 2011; 1(14). http://www.bio-protocol.org/e97",
"DOI": null,
"article": 117
},
{
"id": 3797,
"serial_number": 25,
"pmc": null,
"reference": "Rahman MM, Basaglia M, Favaro L, Boz B, Gumiero B, Casella S. A wooded riparian strip set up for nitrogen removal can affect the water flux microbial composition. Italian Journal of Agronomy.2014; 9(1): 33–37.",
"DOI": null,
"article": 117
},
{
"id": 3798,
"serial_number": 26,
"pmc": null,
"reference": "Rahman MM, Basaglia M, Vendramin E, Boz B, Fontana F, Gumiero B, Casella S. Bacterial diversity of a wooded riparian strip soil specifically designed for enhancing denitrification process. Biology and Fertility of Soils. 2014; 50: 25–35.",
"DOI": null,
"article": 117
},
{
"id": 3799,
"serial_number": 27,
"pmc": null,
"reference": "Marchesi JR, Sato T, Weightman AJ, Martin TA, Fry JC, Hiom SJ, Wade WG. Design and evaluation of useful bacterium-specific PCR primers that amplify genes coding for bacterial 16S rRNA. Applied and Environmental Microbiology.1998; 64:795–799.",
"DOI": null,
"article": 117
},
{
"id": 3800,
"serial_number": 28,
"pmc": null,
"reference": "Osborn M, Moore ERB, Timmis KN. An evaluation of terminal-restriction fragment length polymorphism (T-RFLP) analysis for the study of microbial community structure and dynamics. Environmental Microbiology. 2000; 2(1): 39–50.",
"DOI": null,
"article": 117
},
{
"id": 3801,
"serial_number": 29,
"pmc": null,
"reference": "Lim SM, Yoon MY, Choi GJ, Choi YH. et al. Diffusible and Volatile Antifungal Compounds Produced by an Antagonistic Bacillus velezensis G341 against Various Phytopathogenic Fungi. Plant Pathology Journal. 2017; 33(5): 488–498.",
"DOI": null,
"article": 117
},
{
"id": 3802,
"serial_number": 30,
"pmc": null,
"reference": "Lee T, Park D, Kim K, Lim SM, Yu NH, Kim S, Kim HY. et al. Characterization of Bacillus amyloliquefaciens DA12 showing potent antifungal activity against mycotoxigenic Fusarium species. Plant Pathology Journal. 2017; 33:499–507.",
"DOI": null,
"article": 117
},
{
"id": 3803,
"serial_number": 31,
"pmc": null,
"reference": "Lynch JM, Benedetti A, Insam H. et al. Microbial diversity in soil: ecological theories, the contribution of molecular techniques and the impact of transgenic plants and transgenic microorganisms. Biology and Fertility of Soils. 2004; 40:363–385.",
"DOI": null,
"article": 117
},
{
"id": 3804,
"serial_number": 32,
"pmc": null,
"reference": "Rahman MM, Sultana T, Ali MY, Rahman A. Chemical composition and antibacterial activity of the essential oil and various extracts from Cassia sophera L. against Bacillus sp. from soil. Arabian Journal of Chemistry. 2017; 10(2):2132–2137.",
"DOI": null,
"article": 117
},
{
"id": 3805,
"serial_number": 33,
"pmc": null,
"reference": "Dash BK, Rahman MM, Sarker PK. Molecular Identification of a Newly Isolated Bacillus subtilis BI19 and optimization of production conditions for enhanced production of extracellular amylase. BioMed Research International. 2015; 2015:859805.",
"DOI": null,
"article": 117
},
{
"id": 3806,
"serial_number": 34,
"pmc": null,
"reference": "Burhan A, Nisa U, G ̈okhan C, Omer C, Ashabil A, Osman G. Enzymatic properties of a novel thermostable, ther-mophilic, alkaline and chelator resistant amylase from an alka-liphilic Bacillus sp. isolate ANT-6. Process Biochemistry. 2003; 38: 1397–1403.",
"DOI": null,
"article": 117
},
{
"id": 3807,
"serial_number": 35,
"pmc": null,
"reference": "Adesina MF, Lembke A, Costa R, Speksnijder A, Smalla K. Screening of bacterial activity towards Rhizoctonia solani and Fusarium oxysporum: Site-dependent composition and diversity revealed. Soil Biology and Biochemistry. 2007; 39(11):2818–2828.",
"DOI": null,
"article": 117
},
{
"id": 3808,
"serial_number": 36,
"pmc": null,
"reference": "Ntushelo K, Ledwaba LK, Rauwane ME, Adebo OA, Njobeh PB. The Mode of Action of Bacillus Species against Fusarium graminearum, tools for investigation, and future prospects. Toxins (Basel). 2019;11(10):606.",
"DOI": null,
"article": 117
},
{
"id": 3809,
"serial_number": 37,
"pmc": null,
"reference": "Khan N, Martínez-Hidalgo P, Ice TA, et al. Antifungal activity of Bacillus species against Fusarium and analysis of the potential mechanisms used in biocontrol. Frontiers in Microbiology. 2018; 9:2363.",
"DOI": null,
"article": 117
},
{
"id": 3810,
"serial_number": 38,
"pmc": null,
"reference": "Yuan J, Raza W, Shen Q, Huang Q. Antifungal activity of Bacillus amyloliquefaciens NJN-6 volatile compounds against Fusarium oxysporum f. sp. cubense. Applied and Environmental Microbiology. 2012;78(16):5942-5944.",
"DOI": null,
"article": 117
},
{
"id": 3811,
"serial_number": 39,
"pmc": null,
"reference": "Linhares I, Raposo T, Rodrigues A, Almeida A. Frequency and antimicrobial resistance patterns of bacteria implicated in community urinary tract infections: a ten-year surveillance study (2000-2009). BMC Infectious Diseases. 2013;13: 19",
"DOI": null,
"article": 117
},
{
"id": 3812,
"serial_number": 40,
"pmc": null,
"reference": "Godebo G. Kibru G, Tassew H. Multidrug-resistant bacterial isolates in infected wounds at Jimma University Specialized Hospital, Ethiopia. Annals of Clinical Microbiology and Antimicrobials. 2013; 12: 17.",
"DOI": null,
"article": 117
},
{
"id": 3813,
"serial_number": 41,
"pmc": null,
"reference": "Rana A, Saharan B, Kabi SR, Prasanna R, Sain L. Providencia, a PGPR with biocontrol potential elicits defense enzymes in wheat. Annals of Plant Protection Sciences. 2011; 19(1) :138–141.",
"DOI": null,
"article": 117
}
]
},
{
"id": 114,
"slug": "178-1596045892-phytochemical-constituency-profiling-and-antimicrobial-activity-screening-of-seaweed-extracts-collected-from-the-bay-of-bengal-sea-coasts",
"featured": false,
"slider": false,
"issue": "Vol4 Issue1",
"type": "original_article",
"manuscript_id": "178-1596045892",
"recieved": "2020-07-29",
"revised": null,
"accepted": "2020-10-01",
"published": "2020-10-08",
"pdf_file": "https://jabet.bsmiab.org/media/pdf_file/2023/26/178-1596045892.pdf",
"title": "Phytochemical constituency profiling and antimicrobial activity screening of seaweed extracts collected from the Bay of Bengal sea coasts",
"abstract": "<p>Seaweeds are able to produce a great variety of secondary metabolites that are characterized by a broad spectrum of biological activities. Two seaweeds species, namely <em>Hypnea musciformis </em>and <em>Enteromorpha intestinalis </em>were studied to evaluate the phytochemical constituency and antimicrobial activities. First of all, crude extracts of both sea weeds were prepared by two different extraction methods (soaking and water bath) using different solvents. Phytochemicals profiling results revealed the presence of bioactive compounds (flavonoids, alkaloids, tannin, saponin and phenols) in both seaweed extracts. Quantification results for ethanolic extracts of <em>H. musciformis and E. intestinalis </em>estimated 51 mg and 43 mg tannins in per gram of dried samples and flavonoids contents were found 67 mg and 57 mg/g (Quercetin equivalents) QE/g, respectively. Total phenolic contents were determined in terms of gallic acid equivalent (GAE). <em>H. musciformis </em>exhibited higher amount of phenolics (59 ± 0.0002 mg GAE/g) than <em>E. intestinalis </em>extracts (41 ± 0.0002 mg GAE/g).In antimicrobial activity test, ethanol extracts<em> of H. musciformis </em>and <em>E. intestinalis </em>were found 10 mm of inhibition diameter against all of the bacterial strains. Besides, methanol extracts of <em>E. intestinalis </em>were more susceptible to <em>Staphylococcus aureus and </em><em>Pseudomonas</em> which was close to the inhibition diameter (>15 mm) of the mainstream antibiotic, gentamicin. Moreover, <em>Klebsiella sp. </em>was found more susceptible to ethanol and methanol extracts of <em>H. musciformis </em>as it showed inhibition zone greater than 15 mm. Both Seaweed extracts possessed higher amount of phytochemicals and showed promising antimicrobial activities when compared with the standards.</p>",
"journal_reference": "J Adv Biotechnol Exp Ther. 2021; 4(1): 25-34.",
"academic_editor": "Md Jamal Uddin, PhD; Ewha Womans University, Seoul, South Korea",
"cite_info": "Imran MAS, Bhuiyan FR, et al. Phytochemical constituency profiling and antimicrobial activity screening of seaweed extracts collected from the Bay of Bengal sea coasts. J Adv Biotechnol Exp Ther. 2021; 4(1): 25-34.",
"keywords": [
"Antimicrobial activity",
"Phytochemical",
"Enteromorpha intestinalis",
"Hypnea musciformis",
"Seaweeds"
],
"DOI": "10.5455/jabet.2021.d103",
"sections": [
{
"section_number": 1,
"section_title": "INTRODUCTION",
"body": "<p>Millions of people are being afflicted by different infectious diseases induced by pathogenic bacteria. The phenomena of high mortality rate and the emergence of new multi-drug resistance, bacterial strains have become one of the threatening health problems worldwide [<a href=\"#r-1\">1,2</a>]. Different drug molecules are being applied to combat against those microorganisms. Antibiotics, metal ions, and various quaternary ammonium compounds are being used worldwide though these antimicrobial compounds are also being claimed to be associated with antibiotic resistance, complex chemical synthesis, environmental pollution, and high cost [<a href=\"#r-3\">3,4</a>]. However, natural antibacterial agents have been recently identified to overcome these disadvantages [<a href=\"#r-5\">5</a>]. Marine algae are one of the richest sources of bioactive compounds and chemical diversity [<a href=\"#r-6\">6,7</a>] among them different seaweeds have historical contribution against the predators defense [<a href=\"#r-8\">8,9</a>], and could be a promising antibacterial agents [<a href=\"#r-10\">10,11</a>]. Moreover, algae produce the largest amount of biomass in the marine environmental [<a href=\"#r-12\">12</a>].Seaweeds are plant like ocean organisms that are botanically classified as microphysics marine algae where edible seaweeds are often called sea vegetables, seaweeds come in an amazing variety of beautiful shapes, colors and sizes and found in all of the world’s oceans. They are source materials for structurally unique natural products with pharmacological and biological activities [<a href=\"#r-13\">13</a>]. Among the marine organisms, seaweeds occupy an important place as a source of biomedical compounds [<a href=\"#r-14\">14</a>] and the most interesting algae group because of their broad spectrum of biological activities such as antimicrobial [<a href=\"#r-15\">15</a>], antiviral [<a href=\"#r-16\">16</a>], anti-allergic [<a href=\"#r-17\">17</a>], anticoagulant [<a href=\"#r-18\">18</a>], anticancer [<a href=\"#r-19\">19</a>], antifouling [<a href=\"#r-20\">20</a>] and antioxidant activities [<a href=\"#r-21\">21</a>]. Importantly, seaweeds represent a potential source of antimicrobial substances due to their diversity of secondary metabolites with antiviral, antibacterial and antifungal activities [<a href=\"#r-22\">22</a>]. Structurally diverse secondary metabolites of seaweeds offer defense against herbivores, fouling organisms and pathogens and also play role in reproduction, protection from UV radiation and as allopathic agents [<a href=\"#r-23\">23</a>]. The bactericidal agents found in algae include amino acids, terpenoids, phlorotannins, acrylic acid, phenolic compounds, steroids, halogenated ketones and alkanes, cyclicpolysulphides and fatty acids [<a href=\"#r-24\">24</a>]. Some of these metabolites extracted from seaweed such as iodine, carotene, glycerol, alginates, and carrageenans have been used in pharmaceutical industries [<a href=\"#r-25\">25,26</a>].<br />\r\nThe Bay of Bengal is the northeastern part of the Indian Ocean, bounded on the west and northwest by India on the north by Bangladesh. In Bangladesh, potential seaweeds are being reported in terms of food staffs and pharmaceutical agents from the south-eastern part of the mainland and offshore island [<a href=\"#r-27\">27,28</a>]. Moreover, Bangladesh is rich with around 133 species of seaweeds and eight of them are commercially important. <em>Caulerpa racemosa, Enteromorpha sp, Gelidiella tenuissima, Gelidium pusillum, Halymenia discoidea, Hypnea pannosa, Hydroclathrus clathratus and Sargassum sp </em>are are the commercially important seaweeds in Bangladesh which usually provide low-cost, whole some nutrition and therapeutic protection [<a href=\"#r-29\">29</a>]. Seaweeds may produce a wide variety of secondary metabolites which are characterized by a wide range of biological activities. Still now, there are scanty information regarding the potentiality of these species. This evaluation of <em>H. musciformis</em>, <em>E. intestinalis </em>extracts directly covers the medicinal properties against different pathogenic bacterial organism. That is why the objectives of the present study were toevaluate antimicrobial activity and phytochemical profiling of two widely available seaweed species collected from the Bay of Bengal sea coast.</p>"
},
{
"section_number": 2,
"section_title": "MATERIALS AND METHODS",
"body": "<p><strong>Seaweed material and description of study area</strong><br />\r\nTwo seaweeds species;<em> H. musciformis </em>and <em>E. intestinalis </em>were used in this study. They were freshly collected from North Nuniar Chor, Cox’s bazar (21°35′0″N 92°01′0″E) (<a href=\"#figure1\">Figure 1</a> and <a href=\"#figure2\">2</a>). Collected samples were washed in running water for 10 minutes and then transported to the laboratory and shade dried at 35±3 °C for 36 h. The shade dried seaweeds were preliminarily powdered using electronic blender machine and used for further experiments.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Preparation of seaweed extracts and fractions</strong><br />\r\nThe extracts were prepared by two different methods such as water bath and soaking method [<a href=\"#r-30\">30,31,32</a>]. In both cases, each of 20 grams of powdered seaweed samples were soaked in a conical flask containing 160 ml of the distilled water and different other solvents such as ethanol, methanol and acetone. In soaking method, the samples were gently mixed by shaking and left for 72 hours at room temperature. In water bath method, the samples were mixed gently using the same solvents and left at 65°C for 4 hours. After the soaking at room temperature or by using water bath at specific temperature, the liquid phase was filtered with whatman no. 1 filter paper and allowed to be concentrated at reduced pressure to give specific solvent extracts. The crude extracts were used to test qualitative and quantitative analysis for secondary metabolites and antimicrobial activities against different bacterial strains.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Preliminary qualitative phytochemical analysis</strong><br />\r\nThe presences of secondary metabolites are usually preliminarily traced by the different qualitative phytochemical analysis. Here, several tests had also been conducted for identifying the secondary metabolites present in the alcoholic extracts of <em>H. musciformis </em>and <em>E. intestinalis</em>. The Sodium hydroxide test and the shinoda test were conducted for screening of flavonoids. After the addition of 2 ml of 10% aqueous sodium hydroxide solution into the crude extracts, yellow color precipitation indicated the presence of flavonoid. When yellow color turned into colorless, diluted hydrochloric acid was added. In case of shinoda test, 2 drops of conc. HCl followed by a few fragments of magnesium ribbon were added. Formation of pink, reddish or brown color usually specifies the presence of flavonoids [<a href=\"#r-33\">33</a>]. Few drops of Wagner’s reagents or Dragendorff’s reagent in crude extracts was used for the confirmation of the presence of alkaloids. Appearance of a reddish-brown or orange red precipitation was considered as the positive test for alkaloids [<a href=\"#r-33\">33-36</a>]. Addition of 2-3 drops of 5% ferric chloride to the crude extracts turned into brownish green or a blue-black color results the presence of tannin [<a href=\"#r-35\">35</a>]. A fresh mixture was produced from equal amount of 1% ferric chloride solution and 1% potassium ferrocyanide, 3 drops of the mixture was further added to the extract and filtered this solution, presence of phenol obtained from the formation of a bluish-green color [<a href=\"#r-36\">36</a>]. For the saponin test, 2.5 ml of extract was added to 10 ml of sterile distilled water in a test tube, and then test tube was sealed with cap and shaken vigorously for about 30 seconds. It was then allowed to stand for 30 minutes; formation of honeycomb froth indicated the presence of saponins [<a href=\"#r-35\">35,36</a>].</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Quantitative analysis of phytochemical constituency</strong><br />\r\nQuantitative analyses for phytochemicals were performed for flavonoids, tannin, and phenolics.</p>\r\n\r\n<p> </p>\r\n\r\n<p><em>Determination of total flavonoids content</em><br />\r\nTotal flavonoids content was determined by following the method by Wang [<a href=\"#r-37\">37</a>]. About 0.2 ml of 10% AlCl3, 0.2ml of 1M potassium acetate and 5.6 ml of distilled water followed by 1 ml of methanolic extract at a concentration of 1 mg/ml was added in a test tube to estimate flavonoid content. Blank was prepared without methanol. A series of standard (20, 40, 60,80, 100 µg/ml) was prepared using Quercetin followed by 0.2 ml of AlCl3, 2ml of 1M potassium acetate, 5.6 ml of distilled water. The mixtures were kept at room temperature for 30 minutes. Absorbance was taken at 415 nm using UV-vis spectrophotometer. The result was expressed in mg QE/g of the dried plant extractives.</p>\r\n\r\n<p> </p>\r\n\r\n<p><em>Determination of total tannins content</em><br />\r\nThe tannins content was estimated by Folin-Ciocaltue’s method for both seaweed species as described by Kavita and Indira [<a href=\"#r-37\">37,38</a>]. About 0.1 ml of the sample extract was added to a test tube containing 7.5 ml of distilled water and 0.5 ml of Folin-Ciocaltue phenol reagent. Approximately 1 ml of 35% Na2CO3 diluted to 10 ml with distilled water. The mixtures were shaken well and kept at room temperature for 30 minutes. A set of reference standard solutions of tannic acid (20, 40, 60, 80, 100 µg/ml were prepared. Absorbance for test and standard solutions were measured against the blank at 700 nm with an UV/ Visible spectrophotometer. The estimation of the tannin content was carried out in triplicate. The tannin content was expressed in terms of mg of tannic acid equivalent per g of dried sample.</p>\r\n\r\n<p> </p>\r\n\r\n<p><em>Determination of total phenolics content</em><br />\r\nThe total phenolic content of dry extract was performed with Folin-Ciacaltue assay with slight described by Singleton et al., (1999) with slight modification [<a href=\"#r-36\">36,37</a>]. About 1 ml of methanolic extract sample (1mg/ml) was mixed with 5 ml of 10% Folin Ciaocalteu Reagent and 5 ml of 7.5% Na2CO3. Blank was concomitantly prepared, containing 1 ml methanol, 5 ml of 10% Folin Ciocalteu’s reagent and 5 ml of 7.5% of NaHCO3. The mixtures were incubated for 20 minutes at 25ºC followed by measuring absorbance taken at 760 nm against blank. Gallic acid was used as standard and reactions were performed as triplicates and mean value of absorbance was obtained. Calibration line was constructed using gallic acid as standard. The total content of phenol in extract was expressed in terms of Gallic acid equivalent (mg of GAE/g of extract).</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Antibacterial activity of seaweeds extracts</strong><br />\r\n<em>Microorganisms and media</em><br />\r\nFive bacteria species obtained from the Laboratory of Microbiology, Department of Genetic Engineering and Biotechnology, Shahjalal University of Science and Technology and used as the antimicrobial test strains: There were four gram negative (<em>Escherichia coli, Klebsiella sp., Pseudomonas sp., Salmonella sp</em>.) and one gram positive (<em>Staphylococcus aureus</em>) bacteria. The bacterial strains were maintained on the nutrient agar medium [<a href=\"#r-39\">39</a>].</p>\r\n\r\n<p><em>Agar disk diffusion assay</em><br />\r\nThe screening of antimicrobial activity of the seaweed extracts was carried out with agar disk-diffusion method using Muller Hinton Agar (MHA) medium [<a href=\"#r-34\">34</a>,<a href=\"#r-40\">40</a>]. Bacterial culture (50 µL) was taken from the nutrient broth culture using 100 µL micropipette and poured into the sterile plate containing Muller-Hinton agar medium. Sterile cotton was used for streaking the dried surface of plates. Under aseptic condition, prepared discs (5 mm round filter paper soaked with test solution at a concentration of 1mg/ml) were air dried, placed into center of an agar plate by using a sterile forceps and pressed down. Then discs were employed to be incubated at 37°C within 15 minutes. After 24 & 48 hours of incubation, each plate was examined. There was uniformly circular zone of inhibition on the surface. The diameter of the complete zone of inhibition was measured. All tests were performed in triplicate manner.</p>"
},
{
"section_number": 3,
"section_title": "RESULTS",
"body": "<p><strong>Preliminary qualitative phytochemical analysis</strong><br />\r\nThe present study exposed that ethanolic extracts of <em>H. musciformis </em>and <em>E. intestinalis </em>contained different plant secondary metabolites <em>viz</em> alkaloids, flavonoids, tannins, phenols, and saponins. Preliminary phytochemicals study revealed that <em>E. intestinalis </em>extracts contained flavonoids and saponins in higher extent on the other hand alkaloids and tannins were present in <em>H. musciformis </em>in greater content. Other secondary metabolites were moderately present in two seaweed extracts (<a href=\"#Table-1\">Table 1</a>).</p>\r\n\r\n<div id=\"Table-1\">\r\n<p><a href=\"https://jabet.bsmiab.org/table/178-1596045892-table1/\">Table-1</a><strong>Table 1.</strong> Phytochemical screening of <em>H. musciformis</em> and <em>E. intestinalis</em> collected North Nuniar Chor, Cox’s bazaar, Bangladesh.</p>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Quantitative determination of the </strong><strong>phytochemical</strong><strong> constituency</strong><br />\r\n<a href=\"#Table-2\">Table 2</a> showed the quantitative determination of the tannin, flavonoid and phenolic content. Folin-Ciocaltue’s method was used to determine the total tannin content in different seaweed extracts. The tannin content was expressed in terms of mg of Tannic acid equivalent per g of dried sample. Ethanolic extracts of <em>H. musciformis and E. intestinalis </em>estimated 51 ± 0.0002 and 43 ± 0.0002 mg/g, respectively. However, total flavonoid contents of the extracts were expressed in terms of quercetin equivalent (QE). <em>H. musciformis </em>and <em>E. intestinalis </em>estimated 67 ± 0.0002 and 57 ± 0.0002 mg/g respectively. Total phenolic contents of the extracts were determined in terms of gallic acid equivalent (GAE). <em>H. musciformis </em>exhibited higher amount of phenolics (59 ± 0.0002) present in the crude extracts rather than <em>E. intestinalis </em>extracts (41 ± 0.0002).</p>\r\n\r\n<div id=\"figure1\">\r\n<figure class=\"image\"><img alt=\"\" height=\"269\" src=\"/media/article_images/2024/18/03/178-1596045892-Figure1.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 1.</strong> Sample collection site; North Nuniar Chor, Cox’s bazaar, Banglades (21°35′0″N 92°01′0″E).</figcaption>\r\n</figure>\r\n</div>\r\n\r\n<div id=\"figure2\">\r\n<figure class=\"image\"><img alt=\"\" height=\"226\" src=\"/media/article_images/2024/18/03/178-1596045892-Figure2.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 2. </strong>The pictorial view of <em>Enteromorpha intestinalis</em> and <em>Hypnea musciformis</em> at North Nuniar Chor, Cox’s bazaar, Bangladesh.<strong> </strong></figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<div id=\"Table-2\">\r\n<p><a href=\"https://jabet.bsmiab.org/table/178-1596045892-table2/\">Table-2</a><strong>Table 2. </strong>Quantitative determination of phytochemical constituency.</p>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Antibacterial activity of seaweeds extracts</strong><br />\r\nTwo extraction methods, for instances, soaking and water bath method were applied for generating seaweeds extracts by using ethanol, methanol, acetone and distilled water. The results of antibacterial activities of the seaweeds extracts against selected bacterial strains are summarized in <a href=\"#Table-3\">Table 3</a> in terms of socking method and <a href=\"#Table-4\">Table 4</a> for water batch method.<br />\r\nIn case of soaking method, ethanol extracts of<em> H. musciformis </em>and <em>E. intestinalis </em>were found more than 10 mm inhibition zone against all of the bacterial strains. But, methanol extracts of <em>E. intestinalis </em>were found more active against<em> Staphylococcus aureus </em><em>Pseudomonas</em> as these showed inhibition diameter greater than 15 mm, which was similar to our studied control; Gentamicin (<a href=\"#figure3\">Figure 3</a>). In soaking method ethanol extract of <em>H. musciformis </em>showed highest zone of inhibition (14 ± 0.76 mm) against <em>E. coli </em>and methanol extract of <em>E. intestinalis </em>showed highest zone of inhibition against<em> S. aureus </em>(<a href=\"#figure3\">Figure 3</a>)<em>. </em>Acetone extract of <em>H. musciformis </em>show highest zone of inhibition against <em>S. aureus</em> but which is less than <em>E. intestinalis</em>.<br />\r\nIn case of water bath method, <em>Klebsiella sp. </em>was found more susceptible to ethanol and methanol extracts o<em>f H. musciformis </em>as it showed inhibition zone greater than 15 mm (<a href=\"#figure3\">Figure 3</a>). However, in terms of <em>E. intestinalis, </em>inhibition zone was found more than 10 mm in maximum antimicrobial tests against all bacterial strains. Again, <em>H. musciformis </em>exhibited more active zone of inhibition against different microbes rather than <em>E. intestinalis</em>.</p>\r\n\r\n<div id=\"figure3\">\r\n<figure class=\"image\"><img alt=\"\" height=\"295\" src=\"/media/article_images/2024/18/03/178-1596045892-Figure3.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 3. </strong>Growth inhibition zone of different extracts of <em>H. musciformis</em> and <em>E. intestinalis</em>. (A) <em>E. intestinalis</em> ethanol extract against <em>E. coli </em>(soaking method). (B) <em>E. intestinalis</em> and <em>H. musciformis </em>methanol extract (soaking method) against <em>S. aureus</em>. (C) <em>H. musciformis </em>ethanol extract against <em>Klebsiella </em>sp. (Water bath method). (D) <em>E. intestinalis</em> and <em>H. musciformis</em> acetone extracts against <em>S. aureus </em>(soaking method). (E) Gentamicin against <em>Klebsiella </em>sp. (F) Gentamicin against <em>Staphylococcus aureus.</em></figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<div id=\"Table-3\">\r\n<p><a href=\"https://jabet.bsmiab.org/table/178-1596045892-table3/\">Table-3</a><strong>Table 3.</strong> Screening of antibacterial activity of <em>H. musciformis </em>and <em>E. intestinalis</em> extracts by soaking method.<strong> </strong></p>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<div id=\"Table-4\">\r\n<p><a href=\"https://jabet.bsmiab.org/table/178-1596045892-table4/\">Table-4</a><strong>Table 4. </strong>Screening of antibacterial activity of <em>H. musciformis </em>and <em>E. intestinalis</em> extracts by water bath method.<strong> </strong></p>\r\n\r\n<p> </p>\r\n</div>"
},
{
"section_number": 4,
"section_title": "DISCUSSION",
"body": "<p>Phytochemical profiling of the seaweed samples revealed the presence of different phytochemicals in <em>H. musciformis</em> and <em>E. intestinalis.</em> Phenol, tannin and flavonoids were found in greater amount in both seaweeds extracts. Comparing the quantitative results of both seaweeds extracts it had been found that <em>H. musciformis </em>contained more phytochemicals than <em>E. intestinalis</em>. Moreover, <em>H. musciformis</em> exhibited higher amount of phenolics (59 ± 0.0002 mg/g), tannin (51 ± 0.0002 mg/g) and flavonoid (67 ± 0.0002 mg/g) present in the crude extracts rather than <em>E. intestinalis</em> extracts. These phytochemicals could exhibit antimicrobial activity, Rojas <em>et al</em>. indicated that the antibacterial activity is due to different chemical agents present in the extract such as flavonoids, triterpenoids and other compounds of phenolic nature or free hydroxyl group which was also supported by other study [<a href=\"#r-41\">41, 42, 43</a>]. Extraction methods, different solvents used and time frame of sample collections may influence the antibacterial activity [<a href=\"#r-40\">40</a>]. There were several different organic solvents that had already been suggested to screen algae for antibacterial activity [<a href=\"#r-44\">44</a>]. In the present study, we used two different extraction methods; water bath method and soaking method. Among them <em>H. musciformis </em>showed better results in water bath extraction method and <em>E. intestinalis was better </em>in soaking method. So it could be assumed that <em>H. musciformis </em>release more bioactive compound in water bath method and <em>E. intestinalis</em> in soaking method.<br />\r\nIn current study we used alcoholic and aqueous solvents to generate seaweeds extracts where ethanol extract of <em>E. intestinalis </em>showed highest zone of inhibition against <em>E. coli </em>and methanol extract of <em>E. intestinalis </em>showed utmost zone of inhibition against<em> S. aureus. </em>Ethanol extract of <em>E. intestinalis </em>and methanol extracts of <em>H. musciformis</em> were more effective against selected bacterial strains.<br />\r\nThe seaweed extracts were effective against both gram positive and gram negative bacteria. Kolanjinathan and Stella showed that some macro alge effective against <em>E. coli, P. aeruginosa, S. aureus, K. pneumoniae, E. faecalis </em>[<a href=\"#r-26\">26</a>,<a href=\"#r-45\">45</a>] which strongly supports the study results. There are some other studies which could strengthen our findings, for example, Tuney <em>et al.</em> illustrated that <em>Enteromorpha sp. </em>Is effective against<em> Candida </em>sp.<em>, E. faecalis, S. aureus, S. epidermidis, P. aeruginosa, E. coli </em>[<a href=\"#r-46\">46</a>]. Sukatar <em>et al. </em>reported that <em>Enteromorphalinza </em>is highly susceptible to <em>S. aureus, S. epidermidis, S. fecalis, B. subtilis, S. typhimurium, P. aeruginosa, E. cloacae, E. coli, C. albicans </em>[<a href=\"#r-47\">47</a>].<br />\r\nThe both studied seaweeds, <em>H. musciformis </em>and <em>E. intestinalis </em>could possess potential antimicrobial activities though it needs further comprehensive studies. The availability of secondary metabolites in the natural extracts could directly or indirectly influence the induction of antimicrobial approaches [<a href=\"#r-48\">48,49,50</a>].</p>"
},
{
"section_number": 5,
"section_title": "CONCLUSIONS",
"body": "<p>The priority for the next decades should be focused in the expansion of alternative drugs. Antimicrobial potentiality and phytochemical screening of seaweeds could be the best solution for many pharmaceuticals questions. Much consideration has been funded to the development of innovative projects for the pharmaceutical applications of seaweed, particularly in the design of novel antimicrobial drugs.</p>"
},
{
"section_number": 6,
"section_title": "ACKNOWLEDGMENT",
"body": "<p>We are highly thankful to Professor Dr. Abul Kalam Azad, Department of Genetic Engineering and Biotechnology, Shahjalal University of Science and Technology as he provided us the pathogenic bacterial strains for the study. We are also grateful to Professor Dr. Md. Kamrul Huda, Department of Botany, University of Chittagong for his lab support during the laboratory work of our study. The preprint of the study has already been available at BioRxiv database.</p>"
},
{
"section_number": 7,
"section_title": "AUTHOR CONTRIBUTIONS",
"body": "<p>This work is a collaboration among all the authors. Idea conceptualization, supervision, manuscript finalization and technical support by Mahmudul Hasan. Sample collection, characterization, antibiotic activity screening (Disc Diffusion Method), and manuscript prepared by Md. Abdus Shukur Imran. Sample collection, characterization, antibiotic activity screening (Disc Diffusion Method) by Farhana Rumzum Bhuiyan. Qualitative phytochemical screening by Sheikh Rashel Ahmed and Parsha Shanzana. Tannin by Mahmuda Akter Moli. Flavonoid Screening by Shakhawat Hossain Foysal and Mahmuda Akter Moli. Phenol Screening by Suma Bala Dabi. All authors read and approved the final submitted version of the manuscript.</p>"
},
{
"section_number": 8,
"section_title": "CONFLICTS OF INTEREST",
"body": "<p>There is no conflict of interest among the authors.</p>"
}
],
"figures": [
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/18/03/178-1596045892-Figure1.jpg",
"caption": "Figure 1. Sample collection site; North Nuniar Chor, Cox’s bazaar, Banglades (21°35′0″N 92°01′0″E)",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/18/03/178-1596045892-Figure2.jpg",
"caption": "Figure 2. The pictorial view of Enteromorpha intestinalis and Hypnea musciformis at North Nuniar Chor, Cox’s bazaar, Bangladesh.",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/18/03/178-1596045892-Figure3.jpg",
"caption": "Figure 3. Growth inhibition zone of different extracts of H. musciformis and E. intestinalis. (A) E. intestinalis ethanol extract against E. coli (soaking method). (B) E. intestinalis and H. musciformis methanol extract (soaking method) against S. aureus. (C) H. musciformis ethanol extract against Klebsiella sp. (Water bath method). (D) E. intestinalis and H. musciformis acetone extracts against S. aureus (soaking method). (E) Gentamicin against Klebsiella sp. (F) Gentamicin against Staphylococcus aureus.",
"featured": false
}
],
"authors": [
{
"id": 458,
"affiliation": [
{
"affiliation": "Department of Pharmaceuticals and Industrial Biotechnology, Sylhet Agricultural University, Sylhet-3100, Bangladesh."
}
],
"first_name": "Md. Abdus Shukur",
"family_name": "Imran",
"email": null,
"author_order": 1,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 114
},
{
"id": 462,
"affiliation": [
{
"affiliation": "Department of Botany, University of Chittagong, Chittagong-4331, Bangladesh"
}
],
"first_name": "Farhana Rumzum",
"family_name": "Bhuiyan",
"email": null,
"author_order": 2,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 114
},
{
"id": 463,
"affiliation": [
{
"affiliation": "Department of Plant and Environmental Biotechnology, Sylhet Agricultural University, Sylhet-3100, Bangladesh"
}
],
"first_name": "Sheikh Rashel",
"family_name": "Ahmed",
"email": null,
"author_order": 3,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 114
},
{
"id": 464,
"affiliation": [
{
"affiliation": "Department of Molecular Biology and Genetic Engineering, Sylhet Agricultural University, Sylhet-3100, Bangladesh"
}
],
"first_name": "Parsha",
"family_name": "Shanzana",
"email": null,
"author_order": 4,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 114
},
{
"id": 465,
"affiliation": [
{
"affiliation": "Department of Pharmaceuticals and Industrial Biotechnology, Sylhet Agricultural University, Sylhet-3100, Bangladesh."
}
],
"first_name": "Mahmuda Akter",
"family_name": "Moli",
"email": null,
"author_order": 5,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 114
},
{
"id": 466,
"affiliation": [
{
"affiliation": "Department of Genetic Engineering and Biotechnology, Shahjalal University of Science and Technology, Sylhet-3114, Bangladesh"
}
],
"first_name": "Shakhawat Hossain",
"family_name": "Foysal",
"email": null,
"author_order": 6,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 114
},
{
"id": 467,
"affiliation": [
{
"affiliation": "Department of Pharmaceuticals and Industrial Biotechnology, Sylhet Agricultural University, Sylhet-3100, Bangladesh."
}
],
"first_name": "Suma Bala",
"family_name": "Dabi",
"email": null,
"author_order": 7,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 114
},
{
"id": 468,
"affiliation": [
{
"affiliation": "Department of Pharmaceuticals and Industrial Biotechnology, Sylhet Agricultural University, Sylhet-3100, Bangladesh."
}
],
"first_name": "Mahmudul",
"family_name": "Hasan",
"email": "mhasan.pib@sau.ac.bd",
"author_order": 8,
"ORCID": "https://orcid.org/0000-0003-4761-2111",
"corresponding": true,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "Mahmudul Hasan, Professor; Department of Pharmaceuticals\r\nand Industrial Biotechnology, Faculty of Biotechnology and Genetic Engineering, Sylhet Agricultural University, Sylhet-3100, Bangladesh\r\nE-mail: mhasan.pib@sau.ac.bd",
"article": 114
}
],
"views": 1214,
"downloads": 135,
"references": [
{
"id": 3599,
"serial_number": 1,
"pmc": null,
"reference": "Visvesvara GS, Moura H, Schuster FL. Pathogenic and opportunistic free-living amoebae: Acanthamoeba spp., Balamuthia mandrillaris, Naegleriafowleri, and Sappiniadiploidea. FEMS Immunology & Medical Microbiology. 2007; 50: 1–26.",
"DOI": null,
"article": 114
},
{
"id": 3600,
"serial_number": 2,
"pmc": null,
"reference": "Rose JB, Epstein PR, Lipp EK, Sherman BH, Bernard SM, Patz, JA. Climate variability and change in the United States: potential impacts on water-and foodborne diseases caused by microbiologic agents. Environmental Health Perspectives. 2001; 109(2): 211-221.",
"DOI": null,
"article": 114
},
{
"id": 3601,
"serial_number": 3,
"pmc": null,
"reference": "Levy SB, Marshall B. Antibacterial resistance worldwide: causes, challenges and responses. Nature Medicine. 2002; 12: 122–129.",
"DOI": null,
"article": 114
},
{
"id": 3602,
"serial_number": 4,
"pmc": null,
"reference": "Buffet-Bataillon S, Tattevin P, Bonnaure-Mallet M, Jolivet-Gougeon. AnEmergence of resistance to antibacterial agents: the role of quaternary ammonium compounds-a critical review. International Journal of Antimicrobial Agents. 2012; 5: 381–389.",
"DOI": null,
"article": 114
},
{
"id": 3603,
"serial_number": 5,
"pmc": null,
"reference": "Bernini R, Crisante F, Gentili P, Morana F, Pierini M, Piras M. Chemoselective C-4 aerobic oxidation of catechin derivatives catalyzed by the Trametesvillosa laccase/1-hydroxybenzotriazole system: Synthetic and mechanistic aspects. The Journal of Organic Chemistry. 2011; 76(3): 820-832.",
"DOI": null,
"article": 114
},
{
"id": 3604,
"serial_number": 6,
"pmc": null,
"reference": "Kijjoa A, Sawangwong P. Drugs and cosmetics from the sea. Marine Drugs. 2004; 2: 73-82.",
"DOI": null,
"article": 114
},
{
"id": 3605,
"serial_number": 7,
"pmc": null,
"reference": "Proksch P, Ebel RE, Ebel R. Drugs from the sea-opportunities and obstacles. Marine Drugs. 2003; 1: 5-17.",
"DOI": null,
"article": 114
},
{
"id": 3606,
"serial_number": 8,
"pmc": null,
"reference": "Proksch P. Defensive roles for secondary metabolites from marine sponges and sponge feeding-nudibranchs. Toxicon. 1994; 32: 639-655.",
"DOI": null,
"article": 114
},
{
"id": 3607,
"serial_number": 9,
"pmc": null,
"reference": "Kelecom A, Teixeira VL. Diterpenes of marine brown algae of the family Dictyotaceae: their possible role as defense com-pounds and their use in chemotaxonomy. Science of the Total Environment. 1986; 58(1-2): 109-115.",
"DOI": null,
"article": 114
},
{
"id": 3608,
"serial_number": 10,
"pmc": null,
"reference": "Nygaard K, Lunestad BT, Hektoen H, Berge JA, Hormazabal V. Resistance to oxytetracycline, oxolinic acid and furazolidone in bacteria from marine sediments. Aquaculture. 1992; 104(1-2): 31-36.",
"DOI": null,
"article": 114
},
{
"id": 3609,
"serial_number": 11,
"pmc": null,
"reference": "Vairappan CS, Kawamoto T, Miwa H, Suzuki M. Potent antibacterial activity of halogenated compounds against antibiotic-resistant bacteria. Planta Medica. 2004; 70(11): 1087-1090.",
"DOI": null,
"article": 114
},
{
"id": 3610,
"serial_number": 12,
"pmc": null,
"reference": "Bhadury P, Wright PC. Exploitation of marine algae: biogenic compounds for potential antifouling applications. Planta. 2004. 219(4): 561-578.",
"DOI": null,
"article": 114
},
{
"id": 3611,
"serial_number": 13,
"pmc": null,
"reference": "Faulkner DJ. Marine natural products. Natural Product Reports. 2001; 18(1): 1R-49R.",
"DOI": null,
"article": 114
},
{
"id": 3612,
"serial_number": 14,
"pmc": null,
"reference": "Manilal AS, Sujith J, Selvin GS, Kiran C, Shakir, Lipton AP. Antimicrobial potential of marine organisms collected from the southwest coast of India against multi resistant human and shrimp pathogens. Scientia Marina. 2010; 74(2): 287-296.",
"DOI": null,
"article": 114
},
{
"id": 3613,
"serial_number": 15,
"pmc": null,
"reference": "Zbakh H, Chiheb H, Bouziane H, Sanchez VM, Riadi H. Antibacterial activity of benthic marine algae extracts from the Mediterranean coast of Morocco. Journal of Microbiology, Biotechnology and Food Sciences. 2012; 2(1): 219.",
"DOI": null,
"article": 114
},
{
"id": 3614,
"serial_number": 16,
"pmc": null,
"reference": "Bouhlal R, Haslin C, Chermann JC, Colliec-Jouault S, Sinquin C, Simon G, Bourgougnon N. Antiviral activities of sulfated polysaccharides isolated from Sphaerococcus coronopifolius (Rhodophytha, Gigartinales) and Boergeseniellathuyoides (Rhodophyta, Ceramiales). Marine Drugs. 2011; 9(7): 1187-1209.",
"DOI": null,
"article": 114
},
{
"id": 3615,
"serial_number": 17,
"pmc": null,
"reference": "NA HJ, Moon PD, Lee HJ, Kim HR, Chae HJ, Shin T, Seo Y, Hong SH, Kim HM. Regulatory effect of atopic allergic reaction by Carpopeltisaffinis. Journal of Ethnopharmacology. 2005; 101: 43–48.",
"DOI": null,
"article": 114
},
{
"id": 3616,
"serial_number": 18,
"pmc": null,
"reference": "Dayong S, Jing L, Shuju G, Lijun H. Antithrombotic effect of bromophenol, the alga-derived thrombin inhibitor. Journal of Biotechnology. 2008; 136: 577–588.",
"DOI": null,
"article": 114
},
{
"id": 3617,
"serial_number": 19,
"pmc": null,
"reference": "Kim JY, Lee JA, Kim KN, Yoon WJ, Lee WJ, Park SY. Antioxidative and antimicrobial activities of Sargassummuticum Journal of the Korean Society of Food Science and Nutrition. 2007; 36(6): 663-669.",
"DOI": null,
"article": 114
},
{
"id": 3618,
"serial_number": 20,
"pmc": null,
"reference": "Sunda WK, Kieber DJ, Kiene RP. Huntsman S. An antioxidant function for DMSP and DMS in marine algae. Nature. 2002; 418(6895): 317.",
"DOI": null,
"article": 114
},
{
"id": 3619,
"serial_number": 21,
"pmc": null,
"reference": "Devi GK, Manivannan K, Thirumaran G, Rajathi FAA, Anantharaman P. In vitro antioxidant activities of selected seaweeds from Southeast coast of India. Asian Pacific journal of tropical medicine. 2011; 4: 205–211.",
"DOI": null,
"article": 114
},
{
"id": 3620,
"serial_number": 22,
"pmc": null,
"reference": "Gonzalez del Val, A, Platas G, Basilio A. Screening of antimicrobial activities in red, green and brown macroalgae from Gran Canaria (Canary Islands, Spain). International Microbiology. 2001; 4: 35-40.",
"DOI": null,
"article": 114
},
{
"id": 3621,
"serial_number": 23,
"pmc": null,
"reference": "Watson SB, Cruz-Rivera E. Algal chemical ecology: an introduction to the special issue. Phycologia. 2003; 42(4): 319-323.",
"DOI": null,
"article": 114
},
{
"id": 3622,
"serial_number": 24,
"pmc": null,
"reference": "Aiyegoro OA, Okoh AI. Preliminary phytochemical screening and in vitro antioxidant activities of the aqueous extract of Helichrysumlongifolium BMC Complementary and Alternative Medicine. 2010; 10(1): 21.",
"DOI": null,
"article": 114
},
{
"id": 3623,
"serial_number": 25,
"pmc": null,
"reference": "Kharkwal H, Joshi DD, Panthari PREETI, Pant MKJ, Kharkwal A.C. Algae as future drugs. Asian Journal of Pharmaceutical and Clinical Research. 2012; 5: 1-4.",
"DOI": null,
"article": 114
},
{
"id": 3624,
"serial_number": 26,
"pmc": null,
"reference": "Holdt SL, Kraan S. Bioactive compounds in seaweed: functional food applications and legislation. Journal of Applied Phycology. 2011; 23(3): 543-597.",
"DOI": null,
"article": 114
},
{
"id": 3625,
"serial_number": 27,
"pmc": null,
"reference": "Sarkar MSI, Kamal M, Hasan MM, Hossain MI. Present status of naturally occurring seaweed flora and their utilization in Bangladesh. Research in Agriculture Livestock and Fisheries. 2016; 3(1): 203-216.",
"DOI": null,
"article": 114
},
{
"id": 3626,
"serial_number": 28,
"pmc": null,
"reference": "Sarkar MSI, Kamal M, Hasan MM, Hossain MI, Shikha FH, Rasul MG. Manufacture of different value added seaweed products and their acceptance to consumers. Asian Journal of Medical and Biological Research. 2016; 2(4): 639-645.",
"DOI": null,
"article": 114
},
{
"id": 3627,
"serial_number": 29,
"pmc": null,
"reference": "Ahmed N, Taparhudee W. Seaweed cultivation in Bangladesh: problems and potentials. Journal of Fisheries and Environment. 2005; 28: 13-21.",
"DOI": null,
"article": 114
},
{
"id": 3628,
"serial_number": 30,
"pmc": null,
"reference": "Hwang ES, Do Thi N. Effects of extraction and processing methods on antioxidant compound contents and radical scavenging activities of laver (Porphyratenera). Preventive nutrition and food science. 2014; 19(1):40.",
"DOI": null,
"article": 114
},
{
"id": 3629,
"serial_number": 31,
"pmc": null,
"reference": "Bhuiyan FR, Hasan M, Imran MAS, Ahmed SR, Shanzana P, Marufatuzzahan. Antimicrobial activity screening for three citrus pulp extracts and phytochemical constituency profiling. Journal of Pharmacognosy and Photochemistry. 2019; 8(4): 157-161.",
"DOI": null,
"article": 114
},
{
"id": 3630,
"serial_number": 32,
"pmc": null,
"reference": "Banu VS, Mishra JK. Antimicrobial activity of different solvent based crude extracts from red seaweed Tricleocarpafragilis (L.) Huisman & RA Towns from the coast of South Andaman.The Pharma Innovation Journal. 2018; 7(10): 123-127.",
"DOI": null,
"article": 114
},
{
"id": 3631,
"serial_number": 33,
"pmc": null,
"reference": "Peach K, Tracey MV. Modern methods of plant analysis. Springer Verlag, Berlin, 1956; 3.",
"DOI": null,
"article": 114
},
{
"id": 3632,
"serial_number": 34,
"pmc": null,
"reference": "Bauer AW, Kirby WM, Sherris JC, Turck M. Antibiotic susceptibility testing by a standardized single disk method. American Journal of Clinical Pathology. 1966; 45(4): 493.",
"DOI": null,
"article": 114
},
{
"id": 3633,
"serial_number": 35,
"pmc": null,
"reference": "Harborne JB. Phytochemical methods: A guide to modern techniques of plant analysis, 3rd ed. Chapman & Hall, New York, 1998, pp: 49-188.",
"DOI": null,
"article": 114
},
{
"id": 3656,
"serial_number": 36,
"pmc": null,
"reference": "Trease GE, Evans WC. Pharmacognosy, 15th ed. Saunders Publishers, London, 2002, pp: 42–44, 221–229, 246–249, 304–306, 331–332, 391–393.",
"DOI": null,
"article": 114
},
{
"id": 3657,
"serial_number": 37,
"pmc": null,
"reference": "Kavitha Chandran CI, Indira G. Quantitative estimation of total phenolic, flavonoids, tannin and chlorophyll content of leaves of Strobilantheskunthiana (Neelakurinji). Journal of Medicinal Plants Studies. 2016; 4(4): 282-286.",
"DOI": null,
"article": 114
},
{
"id": 3658,
"serial_number": 38,
"pmc": null,
"reference": "Gupta A, Birhman K, Raheja I, Sharma SK, Kar HK. Quercetin: A wonder bioflavonoid with therapeutic potential in disease management. Asian Pacific Journal of Tropical Disease. 2016; 6(3): 248-252.",
"DOI": null,
"article": 114
},
{
"id": 3659,
"serial_number": 39,
"pmc": null,
"reference": "Singleton VL, Orthofer R, Lamuela-Raventós RM. Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. In Methods in Enzymology. 1999; 299: 152-178.",
"DOI": null,
"article": 114
},
{
"id": 3660,
"serial_number": 40,
"pmc": null,
"reference": "Kandhasamy M, Arunachalam KD. Evaluation of in vitro antibacterial property of seaweeds of southeast coast of India. African Journal of Biotechnology. 2008; 7(12).",
"DOI": null,
"article": 114
},
{
"id": 3661,
"serial_number": 41,
"pmc": null,
"reference": "Marasneh I, Jamal M, Kashasneh M, Zibdeh M. Antibiotic activity of marine algae against multi-antibiotic resistant bacteria. Microbios. 1995; 83(334): 23-26.",
"DOI": null,
"article": 114
},
{
"id": 3662,
"serial_number": 42,
"pmc": null,
"reference": "Cordeiro RA, Gomes VM, Carvalho AF, Melo VM. Effect of proteins from the red seaweed musciformis (Wulfen) Lamouroux on the growth of human pathogen yeasts. Brazilian Archives of Biology and Technology. 2006; 49(6): 915-921.",
"DOI": null,
"article": 114
},
{
"id": 3663,
"serial_number": 43,
"pmc": null,
"reference": "Tuney I, Cadirci BH, Unal D, Sukatar A. Antimicrobial activities of the extracts of marine algae from the coast of Urla (Izmir, Turkey). Turkish Journal of Biology. 2006; 30: 1-5.",
"DOI": null,
"article": 114
},
{
"id": 3664,
"serial_number": 44,
"pmc": null,
"reference": "Sukatar A, Karabay-yavasoglu NU, Ozdemir G, Horzum Z. Antimicrobial activity of volatile component and various extracts of Enteromorphalinza (Linnaeus) J. Agardh from the coast of Izmir, Turky. Annals of Microbiology. 2006; 56(3): 275-279.",
"DOI": null,
"article": 114
},
{
"id": 3665,
"serial_number": 45,
"pmc": null,
"reference": "Moses BJ, Trivedi GK, Mathur HH. Diketosteroid from marine red alga musciformis. Photochemistry. 1989; 28(11): 3237-3239.",
"DOI": null,
"article": 114
},
{
"id": 3666,
"serial_number": 46,
"pmc": null,
"reference": "Ahmed SR, Roy R, Romi IJ, Hasan M, Bhuiyan MKH, Khan MMH. Phytochemical screening, antioxidant and antibacterial activity of some medicinal plants grown in Sylhet region. IOSR Journal of Pharmacy and Biological Sciences. 2019; 14(1): 26-37.",
"DOI": null,
"article": 114
},
{
"id": 3667,
"serial_number": 47,
"pmc": null,
"reference": "Rojas A, Hernandez L, Pereda-Miranda R, Mata R. Screening for antimicrobial activity of crude drug extracts and pure natural products from Mexican medicinal plants. Journal of Ethno pharmacology 1992; 35(3): 275-283.",
"DOI": null,
"article": 114
},
{
"id": 3668,
"serial_number": 48,
"pmc": null,
"reference": "Bisi-Johnson MA, Obi CL, Samuel BB, Eloff JN, Okoh AI. Antibacterial activity of crude extracts of some South African medicinal plants against multidrug resistant etiological agents of diarrhea. BMC complementary and alternative medicine. 2017 Dec 1;17(1):321.",
"DOI": null,
"article": 114
},
{
"id": 3669,
"serial_number": 49,
"pmc": null,
"reference": "Al Akeel R, Al-Sheikh Y, Mateen A, Syed R, Janardhan K, Gupta VC. Evaluation of antibacterial activity of crude protein extracts from seeds of six different medical plants against standard bacterial strains. Saudi journal of biological sciences. 2014 Apr 1;21(2):147-51.",
"DOI": null,
"article": 114
},
{
"id": 3670,
"serial_number": 50,
"pmc": null,
"reference": "Famuyide IM, Aro AO, Fasina FO, Eloff JN, McGaw LJ. Antibacterial activity and mode of action of acetone crude leaf extracts of under-investigated Syzygium and Eugenia (Myrtaceae) species on multidrug resistant porcine diarrhoeagenic Escherichia coli. BMC veterinary research. 2019 Dec;15(1):1-4.",
"DOI": null,
"article": 114
}
]
},
{
"id": 109,
"slug": "178-1597893403-nutritional-status-and-associated-factors-in-children-aged-15-17-years-in-a-suburb-in-vietnam",
"featured": false,
"slider": false,
"issue": "Vol4 Issue1",
"type": "original_article",
"manuscript_id": "178-1597893403",
"recieved": "2020-08-09",
"revised": null,
"accepted": "2020-09-11",
"published": "2020-09-24",
"pdf_file": "https://jabet.bsmiab.org/media/pdf_file/2023/24/178-1597893403.pdf",
"title": "Nutritional status and associated factors in children aged 15-17 years in a suburb in Vietnam",
"abstract": "<p>A double nutritional burden (DBM) has increased rapidly in Vietnam. Finding a solution to reduce the rates of overweight & obesity and thinness was necessary, especially at the age of 15-17 years. This study aimed to investigate nutritional status and elucidate the associated factors in children aged 15-17 years in the suburban of Vietnam. A cross-sectional study was carried out from November to December 2019 in Vietnam. Total of 424 students aged 15-17 years (153 boys and 271 girls) from Hong Thai Highschool using a randomized sampling method. The nutritional status of studied subjects was determined based on the World Health Organization (WHO) criteria of sex and age specific BMI cut-off points for children between 5 and 19 years old (2007). Rates of overweight and obesity was 8.96%, thinness was 8.02%, there was a statistically significant difference between boys’ and girls’ group (p=0.02). Correlation between BMI and WC, HC showed that WC or HC could be another indicator to determine the nutritional status of children. Having milk or a light meal before going to bed was also found to decrease the risk of being overweight by 66.4% (OR = 0.336, 95% CI: 0.116 – 0.972). Eating as much as possible increases the risk of overweight and obesity 2.2 times (p=0.017). Moreover, fast food consumption and physical activity time were no significant difference between the obesity & overweight and non-overweight groups. Especially, compared with the walking group, the group with adults taken had the risk of overweight and obesity 14 times higher, the group traveling by motorbike had the risk of overweight and obesity 5.056 times higher (with p=0.018 and p=0.04, respectively). The results suggest that there was the DBM for children in suburban, nutrition status was not similar among boys and girls. Public health programs should therefore be developed or improved in order to promote good eating habits and physical activity, especially how to move to school among children in Vietnam.</p>",
"journal_reference": "J Adv Biotechnol Exp Ther. 2021; 4(1): 17-24.",
"academic_editor": "Md Jamal Uddin, PhD; Ewha Womans University, Korea",
"cite_info": "Chu DT, Dao DTA, et al. Nutritional status and associated factors in children aged 15-17 years in a suburb in Vietnam. J Adv Biotechnol Exp Ther. 2021; 4(1): 17-24.",
"keywords": [
"Vietnam.",
"Nutritional status",
"Suburban",
"Associated factors",
"Children"
],
"DOI": "10.5455/jabet.2021.d102",
"sections": [
{
"section_number": 1,
"section_title": "INTRODUCTION",
"body": "<p>The childhood nutritional status in the world is currently a prominent issue. In particular, overweight and obesity in children have been a top concern in all areas of the world, not only in high-income countries but also in low-income and middle-income countries [<a href=\"#r-1\">1-3</a>]. The rates of overweight and obesity in developing countries (28.8% overweight, 2.3% obesity) [<a href=\"#r-4\">4</a>] have been similar to high-income countries [<a href=\"#r-5\">5</a>]. In addition to the burden of obesity, the burden of thinness was also popular, creates DBM, especially in developing countries [<a href=\"#r-6\">6, 7</a>]. Southeast Asian Nutrition Survey (SEANUTS) of four countries (Indonesia, Malaysia, Thailand and Vietnam) in 2013 showed that the DBM was differences between four countries [<a href=\"#r-8\">8-12</a>]. In Indonesia, malnutrition was a serious problem with the highest thinness rate among the four countries (39.2% in rural areas and 25.1% in urban areas) [<a href=\"#r-9\">9</a>]. In terms of overweight in urban areas, Vietnam has the highest rate (29%) [<a href=\"#r-12\">12</a>], other side, the highest proportion of rural obesity children in Malaysia (18.1%) [<a href=\"#r-10\">10</a>].<br />\r\nIn Vietnam many years ago, economic reform and poverty reduction (Doi moi) have been implemented. The reform has partly improved the health and nutritional status of Vietnamese people [<a href=\"#r-13\">13</a>]. According to the National Institute of Nutrition statistics in 2018, the prevalence of thinness in Vietnam has reduced from 43% in 2000 to 25% in 2015. However, Vietnam still ranks thirty-six in the world for thinness [<a href=\"#r-13\">13, 14</a>]. Recently, overweight, obesity and nutritionally-related chronic diseases (NRCD) have increased rapidly in Vietnam, mainly in the two big cities of Hanoi and Ho Chi Minh City [<a href=\"#r-12\">12</a>]. Now, Vietnam has to face a DBM [<a href=\"#r-13\">13</a>]. It has posed enormous challenges for policy on the nutrition of Vietnam in the coming period.<br />\r\nMoreover, Vietnam was not much research on nutrition status in children. Most of them have shown that overweight, obesity and thinness have increased and there are differences between rural and urban areas, but was not research on associated factors to the nutritional status [<a href=\"#r-12\">12-14</a>]. A few studies have shown associated factors, for example a study on thinness of children from 0.5 to 15 years old in Ho Chi Minh City in 2019, it showed that the rate of obesity, overweight and thinness were respectively 9.8%, 8.4% and 25.8%. That study also found that thinness in children was significantly associated with breastfeeding during the first hour and obesity was associated with starting complementary foods from equal to/more than 6 months [<a href=\"#r-15\">15</a>]. A study on the risk of obesity in 2017 indicates that birth weight and body mass index of the parents are related to obesity, prenatal weight in the obesity was significantly higher than the non-obesity [<a href=\"#r-16\">16</a>]. However, the above studies point out the associated factors of prenatal nutrition and postnatal care without mentioning the subjects diet factors and active factors, which is one of the factors that directly affect the weight. Other side, almost studies focused on rural and urban areas, there was no research in the suburban area in Vietnam, it was a complex area and gradually attracts many people to live. The majority of studies focused on children under 5 years old or primary and secondary school students, there has been no study to evaluate the nutritional status of high school students (15-17 years old) in Vietnam.<br />\r\nFrom the aforementioned limitation, we conduct research with the aim of evaluating nutritional status and associated factors in children aged 15-17 years in the suburban in Vietnam. Desiring to give an overview and necessary information about the nutritional status of children aged 15-17, thereby contributing to the development of a policy to eliminate the DBM in Vietnam.</p>"
},
{
"section_number": 2,
"section_title": "METHODS AND MATERIALS",
"body": "<p><strong>Study populations</strong><br />\r\nA cross-sectional study was carried out from November to December 2019 Vietnam. We enrolled a total of 424 students (153 boys and 271 girls) using a randomized sampling method from Hong Thai High school in Dan Phuong district, a suburb near Hanoi. Participants’ mean age was 16 years 4 months (ranging from 15 years 11 months to 17 years 11 months), without any significant difference between boys and girls. Pupils who suffered from chronic diseases such as mental disorders, cardiac or metabolic disease related to obesity were not involved in this study.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Measurement</strong><br />\r\nAnthropometric indices of all studied subjects, consisted of body weight, height, waist circumference (WC), and hip circumference (HC), were measured twice and then the mean was computed to use in later analysis. Both participants’ weight and height were measured in the morning when they were standing upright, wearing light clothes without shoes or hats. Figures for weight and height were recorded to the closest 0.5 kg and 1 cm, respectively. Waist circumference was determined by placing a tape measure around the bare stomach, at the middle of the iliac crest and the lower rib margin, while hip was defined as the widest part below the waist. This measurement was separately performed for boys and girls in closed rooms to assure their privacy.<br />\r\nParticipants were asked to fill in a self-administrated questionnaire, which consisted of 2 parts. In part A, besides socio-demographic information, students were required to answer a variety of questions such as yes/no questions about their food hobby or estimate their amount of time spent on physical activities. Meanwhile, part B mostly focused on students’ frequency of eating, each question was designed as a 5-point scale, ranging from “Never” (1 point) to “Always” (5 points). Questions in which higher points related to eating more than normal were categorized into “eating more” group, while the remains were “eating less” questions. All participants were fully explained about the aim of this study as well as how to answer the questionnaire properly.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>BMI calculation and classification of nutritional statues</strong><br />\r\nBody Mass Index (BMI) is a value evaluating body fat based on the weight and height of a person. BMI is calculated as the body weight in kilograms divided by the square of the body height in meter (. The nutritional status of studied subjects was determined based on the World Health Organization (WHO) criteria of sex- and age- specific BMI cut-off points for children between 5 and 19 years old (2007). According to WHO, subjects were classified into 5 different groups: severe thinness, thinness, normal, overweight, and obesity.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Statistical analysis</strong><br />\r\nSPSS version 20.0 (SPSS Inc., Chicago, Illinois) and GraphPad Prism version 8.0 were used to performed analysis. Data in descriptive statistics were expressed as N or Median (interquartile range). The Student’s <em>t-</em>test, nonparametric <em>Mann</em>–<em>Whitney U test, or Fisher’s exact test was used when appropriate. U</em>nivariable logistic regression analysis was also performed to evaluate the odds ratio (OR) of being overweight and obese at 95% significant level. P<0.05 was considered statistically significant.</p>"
},
{
"section_number": 3,
"section_title": "RESULTS",
"body": "<p><strong>Characteristics of studied subjects</strong><br />\r\nThe anthropometric characteristics of studied subjects were given in <a href=\"#Table-1\">Table 1</a>. Of 424 students involved in this study, boys accounted for approximately 36% and there was no difference in age distribution between boy group and girl group. It was obvious that figures for boys in almost all characteristics were considerably higher than for girls and differences were statistically significant, except for hip circumference.</p>\r\n\r\n<div id=\"Table-1\">\r\n<p><a href=\"https://jabet.bsmiab.org/table/178-1597893403-table1/\">Table-1</a><strong>Table 1. </strong>Characteristics of studied subjects with male and female groups. </p>\r\n</div>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Nutritional status of studied subjects</strong><br />\r\nOverall, there were 38 overweight and obese students (accounted for 8.96%), while this figure for thinness and severe thinness was 34 (accounted for 8.02%) (data were not shown). The proportion of obesity and overweight in the boy group was 16.65%, which was considerably higher than in the girl group (only 4.79%). Moreover, the prevalence rate of thinness and severe thinness of boys (1.96% and 9.15%, respectively) were also higher compared with girls (0% and 6.27%, respectively) (<a href=\"#figure1\">Figure 1</a>). Differences were statistically significant.</p>\r\n\r\n<div id=\"figure1\">\r\n<figure class=\"image\"><img alt=\"\" height=\"107\" src=\"/media/article_images/2024/08/03/178-1597893403-Figure1.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 1. </strong>Prevalence of severe thinness, thinness, normal, overweight, and obesity defined by BMI of (A) Total studied subjects, (B) Boy group, and (C) Girl group.</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Association of BMI and other biometric indicators in study subjects</strong><br />\r\nPearson’s correlation coefficient was estimated to analyze the relation between BMI and other body parameters such as weight, height, or waist circumference. <a href=\"#Table-2\">Table 2</a> indicated that BMI was significantly and positively correlated to weight, waist circumference, and hip circumference with high correlation coefficients, which were 0.852, 0.803, and 0.795, respectively. These results suggested that WC or HC could be another indicator to determine the nutritional status of children. Besides, our analysis also revealed that excess belly fat was accompanied by large hip circumference due to their strong correlation with an r=0.829, p<0.001.</p>\r\n\r\n<div id=\"Table-2\">\r\n<p><a href=\"https://jabet.bsmiab.org/table/178-1597893403-table2/\">Table-2</a><strong>Table 2. </strong>Pearson’s correlation analysis for BMI and other biometric parameters in study participants (N=424).</p>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Analysis the association of nutritional status and dietary factors in studied subjects</strong><br />\r\nUnivariate logistic regression was performed (<a href=\"#Table-3\">Table 3</a>) to assess the association between dietary habits and overweight & obesity in students. Our data showed that students of the non-overweight group liked more kinds of food than those of the overweight & obesity group and the difference was statistically significant (p=0.02). This result indicated that consuming various kinds of food may be a protective factor for childhood obesity. Having milk or a light meal before going to bed was also found to decrease the risk of being overweight by 66.4% (OR = 0.336, 95%; CI: 0.116 – 0.972). Interestingly, no significant associations between the frequency of soft drinks, sweets, and fast-food intake per week as well as the average time spending on main meals and obesity were observed.<br />\r\nWhen evaluated the eating habits of students in some specific cases through a 5-point scale questionnaire, we found that obesity and overweight group tended to eat more frequently than the remaining group. In addition, if the mean point of questions related to “eating more than usual” rose by 1, the risk of being overweight and obesity increased more than 2.2 times (p=0.017). Similarly, overweight risk reduced by 45.3% if the mean point of “eating less than usual” questions reduced by 1.</p>\r\n\r\n<div id=\"Table-3\">\r\n<p><a href=\"https://jabet.bsmiab.org/table/178-1597893403-table3/\">Table-3</a><strong>Table 3. </strong>The association of dietary factors and nutritional status between Obesity & overweight group (N=38) and Non-overweight group (N=386). </p>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Analysis the association of nutritional status and active factors in studied subjects</strong><br />\r\nData from <a href=\"#Table-4\">Table 4</a> showed that students of the overweight & obesity group and non-overweight group spend the same amount of daily time on physical activities as well as on screen entertainment. In addition, studied participants tended to adopt a sedentary lifestyle while spending much more time (2-3 hours per day) engaging in an activity like playing videogames, watching TV, or using smartphones than doing physical exercises (0.5-1 hour per day). The proportion of students who loved physical activities or played sports after school was not statistically different between the two groups. However, students’ means of transport were found to be highly associated with obese status. Students who were driven to school by parents increased more than 14 times in the risk of being overweight & obese and the odds ratio for those going to school by motorbike was 5.056 compared to those walking to school (with p=0.018 and p=0.04, respectively).</p>\r\n\r\n<div id=\"Table-4\">\r\n<p><a href=\"https://jabet.bsmiab.org/table/178-1597893403-table4/\">Table-4</a><strong>Table 4. </strong>The association of active factors and nutritional status between Obesity & overweight group (N=38) and Non-overweight group (N=386). </p>\r\n\r\n<p> </p>\r\n</div>"
},
{
"section_number": 4,
"section_title": "DISCUSSION",
"body": "<p>The results showed that the rates of overweight and obesity was 8.96%, thinness was 8.02%, there was a statistically significant difference between boys’ and girls’ group, all proportions of boys group are higher than girls’ group. Correlation between BMI and WC, HC showed that WC or HC could be another indicator to determine the nutritional status of children. Besides, having milk or a light meal before going to bed was also found to decrease the risk of being overweight by 66.4%. Eating as much as possible increases the risk of overweight and obesity 2.2 times. Especially, compared with the walking group, the group with adults taken had the risk of overweight and obesity 14 times higher, the group traveling by motorbike had the risk of overweight and obesity 5.056 times higher.<br />\r\nWe found that the nutritional status of children aged 15-17 years in this study was almost different and lower than other study. Example, a research in Norway about obesity status in 2016 showed that the obesity and overweight rate at the age of 15-17 is 20.1%, 2 times higher than this study (8.96%) [<a href=\"#r-17\">17</a>]. In addition, a research in Hanoi, Vietnam in 2019 found that 17.1% were overweight and 19.1% were obesity at the age of 6-11 [<a href=\"#r-18\">18</a>]. Another study in 2010 on 11-14 year old adolescents in Ho Chi Minh City showed the rates of overweight and obesity were 19.6% and 7.9% [<a href=\"#r-19\">19</a>]. The difference can be explained by the scope of our research was suburban, wasn’t represent of the whole country. Scientific evidence has shown that different levels of economic development have different child nutritional status. Specifically, the rate of overweight in urban areas is higher than in rural areas, and the rate of malnutrition in rural areas is higher than in urban areas [<a href=\"#r-12\">12</a>, <a href=\"#r-20\">20</a>, <a href=\"#r-21\">21</a>]. On the other side, the rates of overweight and obesity of children in suburban seems to be lower than in urban and larger than in rural, rate of thinness is the opposite. Results of the abbreviated SEANUTS study on under 11 years in Vietnam showed that obesity of children account for 29% in urban, while in rural was 5.6% [<a href=\"#r-12\">12</a>]. Compared to the rate of overweight and obesity in suburban in this study (8.96%), we were found that the rate of overweight in suburban area was in the middle of that in rural and urban. In addition, a statistically significant difference between boys and girls group in this study was similar to other studies, almost studies have shown that boys have rate of overweight and obese higher than girls [<a href=\"#r-22\">22</a>]. Interestingly, this study indicates that boys also have significantly higher rates of thinness than girls, this was the special feature of our research. This was an issue that needs to be considered to help balance the nutritional status of the sexes in the future.<br />\r\nIn addition to BMI, WC and HC can be another good indicator to evaluate the nutritional status of children. This was similar to the study in Brazil about children aged 7-10 years, which shows that WC was a good indicator for classifying children’s nutritional status [<a href=\"#r-23\">23</a>]. Two studies in Mexico and China also proved it [<a href=\"#r-22\">22</a>, <a href=\"#r-24\">24</a>], but no studies have yet to prove that HC is also a reliable indicator for classifying nutrition in children, and this was something new in our study. We believe that having milk or light meal was a protective factor from the risk of obesity and overweight. This can be explained by evidence of nocturnal physiology and digestion of food, in particular, the rate of night and day metabolism was equal, so we also needs more energy while sleeping, it can provide energy, help you sleep well, and keeps the next day from starving [<a href=\"#r-25\">25, 26</a>]. On the other hand, fast food consumption and physical activity time were thought have no significant difference between the overweight and obesity and non-overweight groups. It seems to be unsuitable for reality, but it may be due to cross-sectional study, unable to assess causality. Besides, overweight children may have changed their diet, for example, reducing fast food consumption. This can falsify or hide the association between fast food consumption and obesity. Especially, compared with the walking group, the group with adults taken had the risk of overweight and obesity 14 times higher, the group traveling by motorbike had the risk of overweight and obesity 5.056 times higher. Similar to a cross-sectional study of children in Tanzania 2015, the results indicate that children who walk to school were less likely to be overweight than children using transport [<a href=\"#r-27\">27</a>]. As another example, a 2015 study on active committing to school (ACS) in a US state provided evidence of the rate of overweight in children walking to school compared to cycling lower than 7% [<a href=\"#r-28\">28</a>]. Children’s transportation to school has a profound influence on their risk of being overweight and obesity, this evidence was thought to be very helpful in reducing the rate of overweight and obesity children.<br />\r\nThis study has some limitations regarding the generalizability of our findings. First, this study was a cross-sectional study. Therefore, we could not address causal relationships of variables or factors related to nutritional status. The potential causal relationships must be clarified by longitudinal studies. Second, this study only focuses on suburban, the results do not apply to all children in whole country, it was necessary to conduct an extensive study for urban and rural areas to find the difference between each region. In the future, we should conduct more specific study to clarify the associated factors in these findings. Finally, an educational program about daily routines, eating and transportation to school to children and parents was necessary, contributing to reducing the rate of overweight, obesity and thinness based on our study results.</p>"
},
{
"section_number": 5,
"section_title": "ACKNOWLEDGEMENT",
"body": "<p>We would like to thank MSc. Phuong Linh Nguyen (Ban Mai School, Hanoi, Vietnam) for critical reading and checking to improve the manuscript. No funding was received in this study.</p>"
},
{
"section_number": 6,
"section_title": "CONFLICT OF INTEREST",
"body": "<p>All authors have no conflicts of interest or financial ties to disclose.</p>"
},
{
"section_number": 7,
"section_title": "AUTHOR CONTRIBUTIONS",
"body": "<p>DTC, LTD, DTAD, and TNM designed and performed experiments and collected data and informed consents. DTC, VTH, BNL, VTQ, DTT and TNM analysed and interpreted the results, and edited and corrected the manuscript. DTC, VTH, BNL, and VTQ, wrote the manuscript. All authors approved the final manuscript.</p>"
}
],
"figures": [
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/08/03/178-1597893403-Figure1.jpg",
"caption": "Figure 1. Prevalence of severe thinness, thinness, normal, overweight, and obesity defined by BMI of (A) Total studied subjects, (B) Boy group, and (C) Girl group.",
"featured": false
}
],
"authors": [
{
"id": 428,
"affiliation": [
{
"affiliation": "Faculty of Biology, Hanoi National University of Education, 136 Xuan Thuy Road, Cau Giay District, Ha Noi, Vietnam."
}
],
"first_name": "Dinh-Toi",
"family_name": "Chu",
"email": "chudinhtoi.hnue@gmail.com",
"author_order": 1,
"ORCID": null,
"corresponding": true,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "Dinh-Toi Chu, PhD; Faculty of Biology, Hanoi National University of Education, Hanoi, Vietnam. Email: chudinhtoi.hnue@gmail.com",
"article": 109
},
{
"id": 429,
"affiliation": [
{
"affiliation": "Faculty of Biology, Hanoi National University of Education, 136 Xuan Thuy Road, Cau Giay District, Ha Noi, Vietnam."
}
],
"first_name": "Duong Thi Anh",
"family_name": "Dao",
"email": null,
"author_order": 2,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 109
},
{
"id": 430,
"affiliation": [
{
"affiliation": "Faculty of Biology, Hanoi National University of Education, 136 Xuan Thuy Road, Cau Giay District, Ha Noi, Vietnam."
}
],
"first_name": "Le Thi",
"family_name": "Duyen",
"email": null,
"author_order": 3,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 109
},
{
"id": 431,
"affiliation": [
{
"affiliation": "Faculty of Clinical Pharmacy, Hanoi University of Pharmacy, Hanoi, Vietnam"
}
],
"first_name": "Bui Nhat",
"family_name": "Le",
"email": null,
"author_order": 4,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 109
},
{
"id": 432,
"affiliation": [
{
"affiliation": "Institute of Preventive Medicine and Public Health, Hanoi Medical University, Hanoi, Vietnam"
}
],
"first_name": "Vu Thi",
"family_name": "Hue",
"email": null,
"author_order": 5,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 109
},
{
"id": 434,
"affiliation": [
{
"affiliation": "Department of Nutrition, Le Huu Trac National Institute of Burns, Vietnam Military Medical University, Hanoi, Vietnam"
}
],
"first_name": "Vu Tri",
"family_name": "Quang",
"email": null,
"author_order": 6,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 109
},
{
"id": 433,
"affiliation": [
{
"affiliation": "Department of Anatomy, Vietnam Military Medical University, Hanoi, Vietnam"
}
],
"first_name": "Dang Tien",
"family_name": "Truong",
"email": null,
"author_order": 7,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 109
},
{
"id": 435,
"affiliation": [
{
"affiliation": "Department Obstetrics and Gynecology, Hanoi Medical University, Hanoi, Vietnam"
}
],
"first_name": "Thang Nguyen",
"family_name": "Manh",
"email": null,
"author_order": 8,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 109
}
],
"views": 739,
"downloads": 144,
"references": [
{
"id": 3429,
"serial_number": 1,
"pmc": null,
"reference": "Ford ND, Patel SA, Narayan KM. Obesity in Low- and Middle-Income Countries: Burden, Drivers, and Emerging Challenges. Annu Rev Public Health. 2017;38:145-64.",
"DOI": null,
"article": 109
},
{
"id": 3430,
"serial_number": 2,
"pmc": null,
"reference": "Jaacks LM, Kavle J, Perry A, Nyaku A. Programming maternal and child overweight and obesity in the context of undernutrition: current evidence and key considerations for low- and middle-income countries. Public Health Nutr. 2017;20:1286-96.",
"DOI": null,
"article": 109
},
{
"id": 3431,
"serial_number": 3,
"pmc": null,
"reference": "Williams EP, Mesidor M, Winters K, Dubbert PM, Wyatt SB. Overweight and Obesity: Prevalence, Consequences, and Causes of a Growing Public Health Problem. Curr Obes Rep. 2015;4:363-70.",
"DOI": null,
"article": 109
},
{
"id": 3432,
"serial_number": 4,
"pmc": null,
"reference": "Poobalan A, Aucott L. Obesity Among Young Adults in Developing Countries: A Systematic Overview. Curr Obes Rep. 2016;5:2-13.",
"DOI": null,
"article": 109
},
{
"id": 3433,
"serial_number": 5,
"pmc": null,
"reference": "Popkin BM, Slining MM. New dynamics in global obesity facing low- and middle-income countries. Obes Rev. 2013;14 Suppl 2:11-20.",
"DOI": null,
"article": 109
},
{
"id": 3434,
"serial_number": 6,
"pmc": null,
"reference": "Black RE, Victora CG, Walker SP, Bhutta ZA, Christian P, de Onis M, et al. Maternal and child undernutrition and overweight in low-income and middle-income countries. Lancet. 2013;382:427-51.",
"DOI": null,
"article": 109
},
{
"id": 3435,
"serial_number": 7,
"pmc": null,
"reference": "Ng M, Fleming T, Robinson M, Thomson B, Graetz N, Margono C, et al. Global, regional, and national prevalence of overweight and obesity in children and adults during 1980-2013: a systematic analysis for the Global Burden of Disease Study 2013. 2014;384:766-81.",
"DOI": null,
"article": 109
},
{
"id": 3436,
"serial_number": 8,
"pmc": null,
"reference": "Schaafsma A, Deurenberg P, Calame W, van den Heuvel EG, van Beusekom C, Hautvast J, et al. Design of the South East Asian Nutrition Survey (SEANUTS): a four-country multistage cluster design study. Br J Nutr. 2013;110 Suppl 3:S2-10.",
"DOI": null,
"article": 109
},
{
"id": 3437,
"serial_number": 9,
"pmc": null,
"reference": "Sandjaja S, Budiman B, Harahap H, Ernawati F, Soekatri M, Widodo Y, et al. Food consumption and nutritional and biochemical status of 0.5-12-year-old Indonesian children: the SEANUTS study. Br J Nutr. 2013;110 Suppl 3:S11-20.",
"DOI": null,
"article": 109
},
{
"id": 3438,
"serial_number": 10,
"pmc": null,
"reference": "Poh BK, Ng BK, Siti Haslinda MD, Nik Shanita S, Wong JE, Budin SB, et al. Nutritional status and dietary intakes of children aged 6 months to 12 years: findings of the Nutrition Survey of Malaysian Children (SEANUTS Malaysia). Br J Nutr. 2013;110 Suppl 3:S21-35.",
"DOI": null,
"article": 109
},
{
"id": 3439,
"serial_number": 11,
"pmc": null,
"reference": "Rojroongwasinkul N, Kijboonchoo K, Wimonpeerapattana W, Purttiponthanee S, Yamborisut U, Boonpraderm A, et al. SEANUTS: the nutritional status and dietary intakes of 0.5-12-year-old Thai children. Br J Nutr. 2013;110 Suppl 3:S36-44.",
"DOI": null,
"article": 109
},
{
"id": 3440,
"serial_number": 12,
"pmc": null,
"reference": "Le NBK, Le TH, Nguyen DVA, Tran TN, Nguyen HC, Thanh DT, et al. Double burden of undernutrition and overnutrition in Vietnam in 2011: results of the SEANUTS study in 0.5-11-year-old children. Br J Nutr. 2013;110 Suppl 3:S45-56.",
"DOI": null,
"article": 109
},
{
"id": 3441,
"serial_number": 13,
"pmc": null,
"reference": "Khan NC, Khoi HH. Double burden of malnutrition: the Vietnamese perspective. Asia Pac J Clin Nutr. 2008;17 Suppl 1:116-8.",
"DOI": null,
"article": 109
},
{
"id": 3442,
"serial_number": 14,
"pmc": null,
"reference": "Khan NC, Le TD, Ngoc TX, Duong PH, Khoi HH. Reduction in childhood malnutrition in Vietnam from 1990 to 2004. Asia Pac J Clin Nutr. 2007;16:274-8.",
"DOI": null,
"article": 109
},
{
"id": 3443,
"serial_number": 15,
"pmc": null,
"reference": "Huynh G, Huynh QHN, Nguyen NHT, Do QT, Tran KV. Malnutrition among 6-59-Month-Old Children at District 2 Hospital, Ho Chi Minh City, Vietnam: Prevalence and Associated Factors. Biomed Res Int. 2019;2019:6921312.",
"DOI": null,
"article": 109
},
{
"id": 3444,
"serial_number": 16,
"pmc": null,
"reference": "Nguyen HTH, Le TT, Duong DTA, Tao Y, Chu DT. Childhood Obesity Is a High-risk Factor for Hypertriglyceridemia: A Case-control Study in Vietnam. Osong Public Health Res Perspect. 2017;8:138-46.",
"DOI": null,
"article": 109
},
{
"id": 3445,
"serial_number": 17,
"pmc": null,
"reference": "Evensen E, Wilsgaard T, Furberg AS, Skeie G. Tracking of overweight and obesity from early childhood to adolescence in a population-based cohort – the Tromso Study, Fit Futures. BMC Pediatr. 2016;16:64.",
"DOI": null,
"article": 109
},
{
"id": 3446,
"serial_number": 18,
"pmc": null,
"reference": "Pham TTP, Matsushita Y, Dinh LTK, Do TV, Nguyen TTT, Bui AT, et al. Prevalence and associated factors of overweight and obesity among schoolchildren in Hanoi, Vietnam. BMC Public Health. 2019;19:1478.",
"DOI": null,
"article": 109
},
{
"id": 3447,
"serial_number": 19,
"pmc": null,
"reference": "Gerhold DL, Liu F, Jiang G, Li Z, Xu J, Lu M, et al. Gene Expression Profile of Adipocyte Differentiation and Its Regulation by Peroxisome Proliferator-Activated Receptor-γ Agonists. Endocrinology. 2002;143:2106-18.",
"DOI": null,
"article": 109
},
{
"id": 3448,
"serial_number": 20,
"pmc": null,
"reference": "Horiuchi Y, Kusama K, Kanha S, Yoshiike N, team Fr. Urban-Rural Differences in Nutritional Status and Dietary Intakes of School-Aged Children in Cambodia. Nutrients. 2018;11.",
"DOI": null,
"article": 109
},
{
"id": 3449,
"serial_number": 21,
"pmc": null,
"reference": "Nurwanti E, Hadi H, Chang JS, Chao JC, Paramashanti BA, Gittelsohn J, et al. Rural-Urban Differences in Dietary Behavior and Obesity: Results of the Riskesdas Study in 10-18-Year-Old Indonesian Children and Adolescents. Nutrients. 2019;11.",
"DOI": null,
"article": 109
},
{
"id": 3450,
"serial_number": 22,
"pmc": null,
"reference": "Hu L, Huang X, You C, Li J, Hong K, Li P, et al. Prevalence of overweight, obesity, abdominal obesity and obesity-related risk factors in southern China. PLoS One. 2017;12:e0183934.",
"DOI": null,
"article": 109
},
{
"id": 3451,
"serial_number": 23,
"pmc": null,
"reference": "Jensen NS, Camargo Tde F, Bergamaschi DP. Body mass index and waist circumference are good indicators for classifying children’s nutritional status. Cien Saude Colet. 2016;21:1175-80.",
"DOI": null,
"article": 109
},
{
"id": 3452,
"serial_number": 24,
"pmc": null,
"reference": "Megchun-Hernandez M, Espinosa-Raya J, Garcia-Parra E, Albavera-Hernandez C, Briones Aranda A. Association of anthropometric indicators to evaluate nutritional status and cardiometabolic risk in Mexican teenagers. Nutr Hosp. 2019;36:1049-54.",
"DOI": null,
"article": 109
},
{
"id": 3453,
"serial_number": 25,
"pmc": null,
"reference": "Schoffelen PF, Westerterp KR. Intra-individual variability and adaptation of overnight- and sleeping metabolic rate. Physiol Behav. 2008;94:158-63.",
"DOI": null,
"article": 109
},
{
"id": 3454,
"serial_number": 26,
"pmc": null,
"reference": "Goldberg GR, Prentice AM, Davies HL, Murgatroyd PR. Overnight and basal metabolic rates in men and women. Eur J Clin Nutr. 1988;42:137-44.",
"DOI": null,
"article": 109
},
{
"id": 3455,
"serial_number": 27,
"pmc": null,
"reference": "Mwaikambo SA, Leyna GH, Killewo J, Simba A, Puoane T. Why are primary school children overweight and obese? A cross sectional study undertaken in Kinondoni district, Dar-es-salaam. BMC Public Health. 2015;15:1269.",
"DOI": null,
"article": 109
},
{
"id": 3456,
"serial_number": 28,
"pmc": null,
"reference": "Mendoza JA, Liu Y. Active commuting to elementary school and adiposity: an observational study. Child Obes. 2014;10:34-41.",
"DOI": null,
"article": 109
}
]
},
{
"id": 102,
"slug": "178-1597656445-pathophysiological-mechanisms-of-disease-severity-in-covid-19-an-update",
"featured": false,
"slider": false,
"issue": "Special Issue",
"type": "review_article",
"manuscript_id": "178-1597656445",
"recieved": "2020-08-03",
"revised": null,
"accepted": "2020-09-15",
"published": "2020-09-15",
"pdf_file": "https://jabet.bsmiab.org/media/pdf_file/2023/03/178-1597656445.pdf",
"title": "Pathophysiological mechanisms of disease severity in COVID-19: An update",
"abstract": "<p>The pandemic of COVID-19 has created a confounding global health crisis with more than 25 million cases and 870,000 deaths worldwide. The novel coronavirus, SARS-CoV-2, exploits the human ACE2 receptor and potential CD147 to invade a plethora of organ systems including cardiovascular, renal, endocrine, nervous, and gastrointestinal. The pathophysiological mechanisms of COVID-19 infection were found to be associated with the direct viral invasion, dysregulated renin-angiotensin-aldosterone system (RAAS), hypoxia, hyperinflammation, cytokine storm, endotheliopathy, and thrombosis. Emerging evidence suggests that the kinin-kallikrein system, iron dysregulation, and complement component C5a anaphylatoxin have roles in disease severity. In critical patients, the effects manifest as acute respiratory distress syndrome (ARDS), lymphopenia, acute kidney injury (AKI), disseminated intravascular coagulation (DIC), hypovolemic shock leading to multiorgan dysfunction syndrome (MODS). In this review, we provided an update on the pathophysiology of COVID-19 with an emphasis on the clinical outcomes in severe patients that will help facilitate a deeper understanding of the disease.</p>",
"journal_reference": "J Adv Biotechnol Exp Ther. 2020; 3(4): 68-78.",
"academic_editor": "Md. Masudur Rahman, PhD; Sylhet Agricultural University, Bangladesh",
"cite_info": "Siam MHB, Abedin SMM, et al. Pathophysiological mechanisms of disease severity in COVID-19: An update. J Adv Biotechnol Exp Ther. 2020; 3(4): 68-78.",
"keywords": [
"SARS-CoV-2",
"COVID-19",
"Pathogenesis",
"Pathophysiology",
"Multiorgan failure"
],
"DOI": "10.5455/jabet.2020.d158",
"sections": [
{
"section_number": 1,
"section_title": "INTRODUCTION",
"body": "<p>In the matter of a decade, another human coronavirus of zoonotic origin emerged and put the world in an unprecedented crisis. Experiences from SARS (severe acute respiratory syndrome) in 2002, and MERS (Middle East respiratory syndrome) in 2012, led to the prediction of the emergence of a new respiratory disease, and COVID-19 (coronavirus disease-2019) prevailed as a pandemic. The severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) is the causative agent for COVID-19, and it is the seventh coronavirus to infect humans [<a href=\"#r-1\">1</a>]. The SARS-CoV-2 is a relatively large enveloped virus with an RNA genome size of about 30kb. Similar to SARS-CoV, the SARS-CoV-2 belongs to the lineage B of betacoronavirus and shares a 79% of genetic sequence similarity with SARS-CoV, and about 50% with MERS-CoV [<a href=\"#r-2\">2</a>].<br />\r\nIn most cases, COVID-19 causes asymptomatic to mild and moderate illness with fever, cough, dyspnea, and loss of smell although the disease can be life-threatening for critical patients [<a href=\"#r-3\">3</a>]. A recent study has identified six categories of COVID-19 patients based on the clinical manifestations, of which three types of “severity” groups were observed [<a href=\"#r-4\">4</a>]. The severe patients appeared to be elderly, overweight, and had pre-existing comorbidities like asthma or lung disease [<a href=\"#r-4\">4</a>]. Similar to SARS and MERS, the COVID-19 causes viral pneumonia and acute respiratory distress syndrome (ARDS). The hyper inflammation and cytokine storm were previously reported in SARS, and similar immune responses were also observed in COVID-19 [<a href=\"#r-5\">5</a>]. The novel coronavirus has claimed over 870,000 lives globally with an estimated infection fatality rate of 0.65% [<a href=\"#r-6\">6</a>]. In contrast, the case fatality due to SARS was roughly 10%, whereas, for MERS, it was as high as 35% [<a href=\"#r-7\">7</a>]. Studies show that SARS-CoV-2 can cause a plethora of clinical symptoms affecting systems such as cardiovascular, renal, endocrine, nervous, and gastrointestinal [<a href=\"#r-8\">8</a>].<br />\r\nThe current understanding of the various clinical outcomes in COVID-19 is yet in its early stage. However, since SARS-CoV-2 utilizes the same human ACE2 receptor as SARS-CoV and a high genetic similarity is present between two genomes, an analogous pattern of immune reaction is expected [<a href=\"#r-9\">9</a>]. Exploring the pathogenesis and the associated pathophysiology of COVID-19 is fundamental as it bears relevance to both in the bench-side research and the bedside clinical applications. In this narrative review, we aimed to summarize the updated understanding and the advances in the molecular pathogenesis and the pathophysiology of COVID-19 with a focus on the clinical outcomes in severe patients, and discussed the course of disease progression in light of the existing literature.</p>"
},
{
"section_number": 2,
"section_title": "ENTRY, ATTACHMENT, AND REPLICATION",
"body": "<p>SARS-CoV-2 primarily invades the lung cells and causes respiratory illness. It follows a typical lifecycle as of other human coronaviruses. Once inside the body, the virus utilizes its spike protein to attach to the human angiotensin-converting enzyme-2 (ACE2) receptor found on the cell surface [<a href=\"#r-10\">10</a>]. Previously in SARS, the viruses predominantly infected the type-I alveolar (AT1) cells, although the majority (83%) of ACE2 is expressed in type-II alveolar (AT2) cells – indicating that AT2 cells could act as a potential reservoir for the viruses [<a href=\"#r-11\">11</a>].<br />\r\nThe ACE2 receptor is a type-I transmembrane protein with an enzymatically active domain expressed on the surface of many tissues such as lungs, arteries, heart, kidney, and intestine [<a href=\"#r-9\">9</a>]. The primary function of hACE2 is to control vasoconstriction and regulate blood pressure by converting angiotensin-II into vasodilator angiotensin (1 to 7) [<a href=\"#r-12\">12</a>]. As for SARS-CoV-2, the interaction with hACE2 remains to be the primary mechanism of entry into cells although there is a report of using basigin (CD147), also known as extracellular matrix metalloproteinase inducer (EMMPRIN), as a novel route of entry into cells [<a href=\"#r-13\">13</a>].<br />\r\nThe virus enters the cell via receptor-mediated endocytosis pathway [<a href=\"#r-10\">10</a>]. The spike (S) protein is of central importance as it interacts with the host receptor and establishes the gateway for viral entry into the cell. The S-glycoprotein is a homotrimer and it has two halves, S1 and S2. The S protein is cleaved at the S1/S2 junction through transmembrane serine protease TMPRSS2; S1 binds to the receptor while S2 mediates fusion with the cell membrane for entry [<a href=\"#r-14\">14</a>]. Inside the endolysosome, PIKfyve, TCP2, and Cathepsin L contribute to the viral entry. It was also shown that the virus could form syncytium independent of exogenous protease facilitating cell-to-cell fusion [<a href=\"#r-10\">10</a>].<br />\r\nFollowing entry, the virus directs its genome translation by way of host ribosomes to produce polyproteins from ORF1a and ORF1b. The viral encoded proteases, 3-chymotrypsin-like protease (3CLpro) and papain-like protease (PLpro), cleave the polyproteins [<a href=\"#r-15\">15</a>]. A multiprotein replicase-transcriptase complex (RTC) is formed from the non-structural proteins where RdRp plays the key role to produce genomic RNAs [<a href=\"#r-16\">16</a>]. After generating a massive amount of copies, the viruses break free the cell’s borderline by activating different cell death programs. The SARS-CoV-2 exerts similar characteristics as of SARS-CoV in destroying the cells as evidenced by its lethality in cultured human cells [<a href=\"#r-17\">17</a>]. Previously, autopsy studies from SARS showed that apoptosis was present in many organs, including the liver and thyroid gland [<a href=\"#r-18\">18</a>]. In sputum sample, the average viral load was recorded as 7.00 × 106 RNA copies per ml with up to 2.35 × 109 copies per ml at maximum. The viral load was found to be the highest on day 4 with a progressive decline, and remained undetectable after day 28 [<a href=\"#r-19\">19</a>].</p>"
},
{
"section_number": 3,
"section_title": "DYSREGULATION OF RAAS AND KININ-KALLIKREIN SYSTEM",
"body": "<p>Experimental studies show that SARS-CoV-2 infection effectively downregulates the expression of ACE2 [20]. Since ACE2 is a key regulator of the RAAS (renin-angiotensin-aldosterone system) pathway, the downregulation of ACE2 can disrupt the balance between ACE/ACE2 and Ang-II/angiotensin-(1–7) [<a href=\"#r-21\">21</a>]. An increased concentration of Ang-II was shown to aggravate lung injury through vascular permeability and pulmonary edema [<a href=\"#r-22\">22,23</a>]. Of note, the RAAS consists of a network of regulatory proteins that operate in important processes, for instance, electrolyte balance, vascular permeability, and blood pressure regulation [<a href=\"#r-21\">21</a>]. The RAAS pathway elevates blood pressure as renin initiates the system by converting angiotensinogen (produced by the liver) into angiotensin-I, and subsequently to angiotensin-II by ACE, acting as a positive regulator of the system to stimulate vasoconstriction. Conversely, the destruction of alveolar cells and downregulation of ACE2 cause overactivation of the RAAS system. The upregulation of Ang-II levels leads to the loss of the protective effects of angiotensin-(1–7) and contributes to severe lung injury, and the resulting damage releases inflammatory mediators leading to pyroptosis [<a href=\"#r-18\">18</a>,<a href=\"#r-24\">24,25</a>].<br />\r\nMoreover, the downregulation of ACE2 expression is hypothesized to have impacted the kinin-kallikrein system. The kallikreins are serine-protease enzymes that convert kininogen into kinin plasma protein. The activation of the kinin-kallikrein system releases bradykinins that bind to its receptor-B2 found on the endothelial cells to initiate relaxation of vascular smooth muscle and increase in vascular permeability [<a href=\"#r-26\">26</a>]. Generally, the ACE and ACE2 work as kininase II enzymes to breakdown bradykinin metabolites; however, the downregulation of ACE2 impairs the balance in the system and increases bradykinin levels. Failure to neutralize the kinins can result in pulmonary injury, and angioedema [<a href=\"#r-27\">27</a>].</p>"
},
{
"section_number": 4,
"section_title": "INFLAMMASOME AND INFLAMMATORY MEDIATORS",
"body": "<p>The SARS-CoV-2 genome contains the viroporin 3a which is a potent activator of cytoplasmic nod-like receptor family pyrin domain-containing 3 (NLRP3) inflammasome as seen in SARS-CoV [<a href=\"#r-28\">28</a>]. A study suggests that proinflammatory cytokines are produced in response to the formation of NLRP3 inflammasome [<a href=\"#r-28\">28</a>]. The inflammasomes are multiprotein complex in the cytosol that promote the secretion of IL-1β, IL-18, and damage-associated molecular patterns (DAMPs). Excessive release of DAMPs due to hyperactivation of NLRP3 inflammasome can result in the release of high mobility group box 1 (HMGB1), infiltration of neutrophils, activation of macrophages, pyroptosis, and cytokine production [<a href=\"#r-29\">29</a>].<br />\r\nStudies show that a significantly elevated level of pro-inflammatory cytokines including IL-2, IL-7, IL-10, granulocyte-macrophage colony-stimulating factor (GM-CSF), interferon gamma-induced protein 10 (IP10), monocyte chemoattractant protein 1 (MCP1), macrophage inflammatory protein 1-alpha (MIP1α), and TNFα was found in patients affected with COVID-19 [<a href=\"#r-30\">30</a>]. The activated CD4<sup>+</sup> T cells, macrophages, dendritic cells, and epithelial cells secrete these cytokines and chemokines in response to inflammation [<a href=\"#r-31\">31–33</a>]. The release of inflammatory mediators brings CD14<sup>+</sup> CD16<sup>+</sup> monocytes into pulmonary circulation where monocytes mature into macrophages and subsequently release notably IL-1. IL-6, TNFα and HMGB1 [<a href=\"#r-8\">8</a>,<a href=\"#r-34\">34,35</a>]. The HMGB1 acts both as a cytokine and a nuclear protein; and plays an important role in endothelial activation, and systemic inflammation [<a href=\"#r-35\">35</a>]. The inflammation recruits natural killer (NK) cells and T cells producing GM-CSF, and tumor necrosis factor (TNF) inviting more macrophages to the site of infection [<a href=\"#r-30\">30</a>].<br />\r\nThe elevated levels of cytokines cause vasodilation and increase the capillary permeability by relaxing the smooth muscle and contracting the endothelial cells. This, in turn, allows blood plasma to leak into interstitial space, and to alveoli to cause alveolar edema. The pyrogenic cytokines travel via the blood to the central nervous system and get the hypothalamus to reset its thermostat to increase the body temperature. The inflammatory mediators, in particular IL-8, bring neutrophil to the scene to destroy the viruses by releasing reactive oxygen species (ROS) and an arsenal of enzymes into the surrounding tissues [<a href=\"#r-36\">36</a>].</p>"
},
{
"section_number": 5,
"section_title": "COLLATERAL DAMAGE: NEUTROPHILS AND HYPERACTIVE T CELLS",
"body": "<p>In COVID-19, laboratory tests found leucopenia with leukocyte counts of 2.91 × 109 cells/L of which 70.0% were neutrophils [<a href=\"#r-30\">30</a>]. Neutrophils are important in viral infection as they keep the inflammation in check. Albeit, unless a balance is maintained between its beneficial and detrimental effects, they harm the host tissue by collateral damage [<a href=\"#r-34\">34</a>]. The autopsy results from people with COVID-19 showed an extensive accumulation of neutrophil in pulmonary capillaries [<a href=\"#r-36\">37</a>].<br />\r\nThe damage of type-I and type-II pneumocytes impairs the gas exchange and surfactant production which inevitably leads to the increase of surface tension. The alveoli collapse and fluid containing proteins build up inside the alveoli. The cellular debris and consolidated proteinaceous fluid stimulate coughing, while the decrease in gas exchange results in hypoxia. An evaluation of the host immune system in COVID-19 critical patients found that the extent of hypoxemia was closely related to the levels of host immune cells [<a href=\"#r-38\">38</a>]. The low partial pressure of oxygen prompts chemoreceptors to stimulate the sympathetic nervous system (SNS) to increase both heart rate and respiration rate; consequently, shortness of breath occurs.<br />\r\nThe progression of the disease is also accounted for the reduced lymphocyte counts and the persistently low level of type-I interferon (IFN) induction [<a href=\"#r-34\">34</a>]. Generally, the type-I IFN plays a crucial role in antiviral response by preventing viral replication and spread. However, it has been shown that beta coronaviruses can sequester their viral RNA from pattern recognition receptors (PRR) while the nonstructural proteins such as nsp3 and nsp1 can block the interferons to bind to its IFN receptors [<a href=\"#r-39\">39,40</a>]. It is speculated that the SARS-CoV-2 infection might not effectively activate IFN-regulatory factor 3 (IRF3) and IRF7 which could contribute to reduced interferon response [<a href=\"#r-39\">39,41</a>].<br />\r\nThe global T cell lymphopenia seen in COVID-19 severe patients is yet to be clarified in detail, but tentative studies suggest that Fas-mediated activation-induced cell death (AICD) is the likely cause of lymphocytopenia and impaired immune response [<a href=\"#r-42\">42</a>]. Moreover, the hyperactivated CD8 T cells with a high proportion of HLA-DR (CD4 3.47%) and CD38 (CD8 39.4%) were observed in COVID-19 [<a href=\"#r-43\">43</a>]. In CD4 T cells, there was an increased concentration of highly proinflammatory CCR6<sup>+</sup> Th17. The cytotoxic T lymphocytes (CTL) were equipped with high concentrations of cytotoxic granules: granzyme A, granzyme B, granzyme K, granzyme M and perforin [<a href=\"#r-44\">44</a>]. The CTLs are supposed to initiate programmed cell death to prevent the survival of the invading virus but hyperreactive CTLs could show high cytotoxicity to induce apoptosis in healthy cells and lead to the subsequent severity of the disease [<a href=\"#r-43\">43</a>].</p>"
},
{
"section_number": 6,
"section_title": "THE COMPLEMENT CASCADES",
"body": "<p>As the first line of defense, the complement system exerts substantial action against various viral attacks [<a href=\"#r-45\">45</a>]. However, in the case of SARS-CoV, one experimental study indicated that instead of protective effects, the complement system could potentially increase the chance for lung damage [<a href=\"#r-46\">46</a>]. The C3a and C5a can recruit inflammatory cells to the site, and the recruitment of large amounts of neutrophil and macrophages can exert pro-inflammatory activity [<a href=\"#r-47\">47</a>]. One study demonstrated that patients who died of ARDS in COVID-19 had an increased amount of CD163<sup>+</sup> macrophages infiltration in the lungs and the macrophages expressed complement component 5a receptor 1 (C5aR1). The study also reported that the soluble level of C5a anaphylatoxin was linked to COVID-19 disease severity [<a href=\"#r-48\">48</a>]. Another study reported that patients who suffered from respiratory failure in COVID-19 had an excessive amount of C5b-9, C4d, and MASP-2 deposition in the microvasculature [<a href=\"#r-49\">49</a>]. Besides, the inflammatory chemokines such as CCL4 (macrophage inflammatory protein-1β), CCL2 (monocyte chemoattractant protein 1) and CXCL9 (monokine induced by gamma interferon) might have important roles in disease severity in COVID-19 [<a href=\"#r-48\">48</a>].</p>"
},
{
"section_number": 7,
"section_title": "IRON DYSREGULATION, GLUTATHIONE, AND FERROPTOSIS",
"body": "<p>Hyperferritinemia has been observed in COVID-19 patients and described as a marker of disease severity [<a href=\"#r-50\">50–52</a>]. Iron plays an important role in oxygen transport from the lungs to the body; and the cells store irons as ferritin in its ferric (Fe<sup>3+</sup>) state. During infection, the interleukin-6 stimulates the synthesis of hepcidin which is a key regulator-hormone of iron, and hepcidin increases the intracellular concentration of ferritin [<a href=\"#r-53\">53,54</a>]. The excess amount of intracellular irons interacts with molecular oxygen and generates ROS which can have damaging effects on the cells [<a href=\"#r-55\">55</a>]. The accumulation of abundant accessible intracellular iron contributes to ferroptosis which is an iron-dependent regulated form of oxidative cell death [<a href=\"#r-56\">56</a>].<br />\r\nInside the cell, the glutathione (GHS) acts as an antioxidant and provides the primary protection against ferroptosis. Depletion of glutathione leads to loss of cellular antioxidant ability, and inhibition of enzymes dependent on glutathione, such as glutathione peroxidases [<a href=\"#r-57\">57</a>]. One study showed that endogenous deficiency of glutathione can result in severe clinical complications in COVID-19 patients [<a href=\"#r-58\">58</a>]. The glutathione deficiency was found to be associated with male sex, age over 65 years, chronic disease, cigarette smoking, and low glutathione intake linked to insufficient consumption of fresh vegetables [<a href=\"#r-58\">58</a>]. One experimental study denoted that GSH deficiency and its related elevated oxidative stress could epigenetically change the expression of regulating genes of vitamin D contributing to a secondary vitamin D deficiency [<a href=\"#r-59\">59</a>]. In summary, the disruption of iron homeostasis and the resulting ferroptosis can affect multiple organ systems including liver, kidney, heart, and lung [<a href=\"#r-60\">60</a>].</p>"
},
{
"section_number": 8,
"section_title": "EXTRAPULMONARY SYMPTOMS",
"body": "<p>A wide variety of clinical manifestations is observed in COVID-19 as the virus infects gastrointestinal, neurologic, renal, and myocardial tissues [<a href=\"#r-8\">8</a>]. The gastrointestinal complications including anorexia, nausea, diarrhea, and abdominal pain could be directly attributed to viral invasion as SARS-CoV-2 was found to productively infect gut enterocytes [<a href=\"#r-61\">61</a>]. Similarly, the manifestations of anosmia, hypogeusia, headache, and visual dysfunction suggest a potential retrograde axonal transport of the virus into the brain or via nasal mucosa, lamina cribrosa and the olfactory nerve [<a href=\"#r-8\">8</a>]. Another study speculated the transport of the virus using motor proteins dynein and kinesin via afferent nerve endings from the lungs [<a href=\"#r-62\">62</a>]. However, one study hypothesized that the disrupted sense of smell could ironically be an evolutionary mechanism to self-destruct olfactory neurons to halt viral tropism into the central nervous system [<a href=\"#r-63\">63</a>]. Since the olfactory neurons regenerate every 30 – 120 days, the resulting anosmia could act as a self-defense mechanism which is supported by studies that indicate: anosmia may be a predictor of good prognosis for COVID-19 [<a href=\"#r-64\">64,65</a>]. Despite, the evidence for viral tropism in neuron cells suggests that circulating leukocytes could alternatively transport SARS-CoV-2 across the blood-brain barrier in a “Trojan horse” manner [<a href=\"#r-66\">66</a>].<br />\r\nHigh expression of ACE2 receptors in myocardial tissue and subsequent isolation of virus from these tissues support the possible mechanism of virus-mediated cardiac dysfunction, resulting in myocardial injury, arrhythmias, cardiomyopathy, and acute coronary syndromes (ACS) [<a href=\"#r-8\">8</a>]. In severe patients, the low partial pressure of oxygen and hyper-inflammation led to the alteration of liver biomarkers and liver damage [<a href=\"#r-67\">67</a>]. The elevated levels of cytokines may lead to apoptosis of pancreatic β-cell and a range of abnormalities such as worsened hyperglycemia and ketoacidosis. Endocrinologic complications were more pronounced in patients with pre-existing diabetes and obesity [<a href=\"#r-8\">8</a>]. In addition, histopathologic studies suggest that the virus could directly infect the renal system damaging the kidney tubule cells; and in combination with hyperimmune reaction, the acute kidney injury (AKI) manifests as it impairs the normal body fluid flow [<a href=\"#r-8\">8</a>]. The health records of 5,449 COVID-19 patients hospitalized in New York city hospitals revealed that up to 36.6% of them developed AKI, in which more than 30% were at stage 3, and often requiring renal replacement therapy (RRT) [<a href=\"#r-68\">68</a>].</p>"
},
{
"section_number": 9,
"section_title": "CYTOKINE STORM AND SYSTEMIC INFLAMMATION",
"body": "<p>In the cytokine storm, the elevated levels of cytokines contribute to vasodilation, systemic inflammation, and immune suppression. In the lungs, the widespread inflammation appears as acute respiratory distress syndrome (ARDS) and in conjunction with cytokines, it may cause systemic inflammatory response for some patients [<a href=\"#r-69\">69</a>]. Respiratory failure could occur due to ARDS which is the likely cause of death in 70% of fatal COVID-19 cases [<a href=\"#r-70\">70</a>]. Aging was found to be correlated with elevated proinflammatory cytokines which might imply why children are less affected in COVID-19 [<a href=\"#r-71\">71</a>].<br />\r\nThe endothelial activation due to the cytokine-release by activated neutrophils and macrophages secrete TNF-α and IL-1β. The formation of neutrophil extracellular traps (NETs) further damages the endothelium and stimulates the release of IL-6, IL-8, nitric oxide (NO), ROS, and platelet-activating factor (PAF) [<a href=\"#r-8\">8</a>,<a href=\"#r-34\">34</a>]. The subsequent vasodilation, increased permeability and decreased perfusion within the systemic circulation allow fluids to leak out and accumulate in the tissues. Blood volume and total peripheral resistance (TFR) decrease – resulting in the fall of blood pressure. With severe hypovolemia and low perfusion, hypovolemic shock takes place.<br />\r\nThe systemic effects of cytokine storms recruit secondary immune mediators such as human cytokine synthesis inhibitory factor (CSIF) or IL-10 that suppresses cytokine production; and soluble tumor necrosis factor receptors (sTNFR) that neutralize circulating TNF-α and induce apoptosis. However, in COVID-19, all three types of immune responses (type-I, II, and III) are triggered and they subdue the buffering capacity of the host to maintain immune balance [<a href=\"#r-72\">72</a>]. In response to high viral load, the IFN-α, IFN-γ, and TNF-α cytokines are overproduced and inflammasome-induced cytokines (i.e. IL-1β and IL-18) rise significantly [<a href=\"#r-72\">72</a>]. One study demonstrated four distinct immune signatures where patients with severe outcomes had shown a sustained elevated cytokine response of all four types, particularly IFN-λ, TGF-α, IL-16, IL-23, IL-33 and coagulopathy markers [<a href=\"#r-72\">72</a>].</p>"
},
{
"section_number": 10,
"section_title": "MULTIPLE ORGAN DYSFUNCTION SYNDROME",
"body": "<p>The inflammation of the endothelium marked by the presence of activated neutrophils and macrophages, and the decreased expression of ACE2 after SARS-CoV-2 infection initiate pro‐inflammatory and procoagulant pathways [<a href=\"#r-73\">73</a>]. The higher expression of tissue factor (TF) and plasminogen activator inhibitor 1 (PAI-1) and the impairment of tissue factor pathway inhibitor (TFPI) allow the formation of fibrin-based blood clots throughout the bloodstream, particularly in the capillaries [<a href=\"#r-34\">34</a>]. One study provided biochemical evidence and showed that endotheliopathy was widespread among critically ill patients of COVID-19 [<a href=\"#r-74\">74</a>].<br />\r\nThe occurrence of thromboinflammation causes disseminated intravascular coagulation (DIC) leading to fibrin deposition and microvascular thrombosis. Study shows that hypoxia-mediated hyperviscosity and the upregulation of the HIF-1 (hypoxia-inducible factor 1) may contribute prothrombotic state [<a href=\"#r-75\">75</a>]. The deposition of microthrombi and vasculitis could manifest as cutaneous manifestation. The thrombi could travel through the circulatory system and create a blockage in the supply of blood and oxygen in a particular organ, resulting in tissue ischemia. The occurrence of pulmonary embolism and deep venous thrombosis (DVT) has been frequently reported in COVID-19 pneumonia patients [<a href=\"#r-76\">76</a>]. Moreover, emerging evidence suggests that thrombosis could take place in intravenous catheters, extracorporeal circuits leading to myocardial infarction, and ischemic stroke [<a href=\"#r-8\">8</a>]. Serum inflammatory markers such as C-reactive protein (CRP), D-dimer, and fibrinogen are prognostic of subsequent severity and mortality in COVID-19 [<a href=\"#r-8\">8</a>,<a href=\"#r-77\">77,78</a>].<br />\r\nWith cytokine storm mediated systemic inflammation, hypoxia, hypotension, and thrombosis, the multi-organ damage takes place especially in the form of cardiac, hepatic, and renal failure, leading to multi-organ dysfunction syndrome (MODS). The physiologic derangement associated with MODS can involve two or more organ systems failure, eventually, progress to an irreversible state and death. <a href=\"#figure1\">Figure 1</a> shows a schematic diagram of COVID-19 pathophysiology leading to multi organ failure.</p>"
},
{
"section_number": 11,
"section_title": "FUTURE PERSPECTIVES",
"body": "<p>With the ability to invade multiple organ systems and disrupt a wide range of pathways, the COVID-19 has presented itself as a complex biological phenomenon that remains to be elucidated in detail. Areas of major interests include the proper understanding of how the downregulation of ACE2 affects the kinin-kallikrein system (KKS) and blood pressure; and how the development of thrombotic complications in COVID-19 is associated with the kinin-kallikrein system.<br />\r\nSince type-I interferons are part of the body’s immune-combat strategy against viruses, the cause for the reduced type-I interferon production in COVID-19 patients is an open area of research. In addition, the evidence for the direct invasion of SARS-CoV-2 in different cell types will uncover the degree and extent of infection. T cell lymphopenia, iron dysregulation, and neural tropism are also among the highlights for further research.<br />\r\nIt is notable to mention that multiple organ dysfunction syndrome (MODS) is suggested to be different from the multisystem inflammatory syndrome in children (MIS-C) [<a href=\"#r-79\">79</a>]. The MIS-C is likely the result of an autoimmune response mediated by antibodies affecting a number of organs; and the syndrome may lead to fatality. The potential for antibody-dependent enhancement (ADE) needs to be investigated since ADE may exert an adverse outcome in vaccination. With an in-depth understanding of pathogenesis and pathophysiology, it will be possible to devise effective strategies to fight the COVID-19.</p>\r\n\r\n<div id=\"figure1\">\r\n<figure class=\"image\"><img alt=\"\" height=\"537\" src=\"/media/article_images/2024/24/07/178-1597656445-Figure1.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 1. </strong>A schematic diagram of COVID-19 pathophysiology. The SARS-CoV-2 binds to the ACE2 receptor and invades multiple organ systems. The dysregulation of the renin–angiotensin–aldosterone system (RAAS) and the Kinin-Kallikrein system contributes to increase permeability and vasodilation. The release of proinflammatory cytokines by white blood cells contributes to widespread inflammation, plasma leakage and fall in blood pressure. The development of ARDS, systemic infection, disseminated intravascular coagulation (DIC) and tissue ischemia lead to multiple organ dysfunction syndrome (MODS).</figcaption>\r\n</figure>\r\n</div>"
},
{
"section_number": 12,
"section_title": "CONCLUSIONS",
"body": "<p>The COVID-19 pandemic has become a global concern as the disease is likely to persist for a long time. The wide range of clinical manifestations beyond the respiratory system warrants rigorous research to develop a comprehensive scenario of COVID-19 disease. topics including the mechanism of direct viral injury, the potential alternative routes of invasion, the role of bradykinins in blood clots, ferroptosis, T cell apoptosis, and nerve cell damage in anosmia require special focus to answer the points at issue. We believe our updated study of COVID-19 pathophysiology will contribute to a holistic understanding of the mechanism of disease development, and thus provide a window for better strategies in therapeutic interventions and patient care.</p>"
},
{
"section_number": 13,
"section_title": "ACKNOWLEDGEMENT",
"body": "<p>This research received no external funding.</p>"
},
{
"section_number": 14,
"section_title": "CONFLICTS OF INTEREST",
"body": "<p>The authors declare no conflict of interest.</p>"
},
{
"section_number": 15,
"section_title": "AUTHOR CONTRIBUTIONS",
"body": "<p>MHB Siam designed the study and wrote the manuscript. MHB Siam, and SMM Abedin carried out the literature review. A Ahmed and MHB Siam performed the illustration. MS Hossain supervised the study. NH Nishat, MS Hossain, and A Ahmed provided critical intellectual contribution in reviewing and revising the manuscript. All authors read and approved the final submitted version of the manuscript.</p>"
}
],
"figures": [
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/24/07/178-1597656445-Figure1.jpg",
"caption": "Figure 1. A schematic diagram of COVID-19 pathophysiology. The SARS-CoV-2 binds to the ACE2 receptor and invades multiple organ systems. The dysregulation of the renin–angiotensin–aldosterone system (RAAS) and the Kinin-Kallikrein system contributes to increase permeability and vasodilation. The release of proinflammatory cytokines by white blood cells contributes to widespread inflammation, plasma leakage and fall in blood pressure. The development of ARDS, systemic infection, disseminated intravascular coagulation (DIC) and tissue ischemia lead to multiple organ dysfunction syndrome (MODS).",
"featured": false
}
],
"authors": [
{
"id": 389,
"affiliation": [
{
"affiliation": "Department of Emerging and Neglected Diseases, Biomedical Research Foundation, Dhaka, Bangladesh"
},
{
"affiliation": "Department of Microbiology, University of Dhaka, Dhaka, Bangladesh"
}
],
"first_name": "Md. Hasanul Banna",
"family_name": "Siam",
"email": null,
"author_order": 1,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 102
},
{
"id": 390,
"affiliation": [
{
"affiliation": "Department of Microbiology, University of Dhaka, Dhaka, Bangladesh"
}
],
"first_name": "Shah MD Minhajul",
"family_name": "Abedin",
"email": null,
"author_order": 2,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 102
},
{
"id": 391,
"affiliation": [
{
"affiliation": "Department of Emerging and Neglected Diseases, Biomedical Research Foundation, Dhaka, Bangladesh"
},
{
"affiliation": "Dhaka Medical College and Hospital, Dhaka, Bangladesh"
}
],
"first_name": "Ahsan",
"family_name": "Ahmed",
"email": null,
"author_order": 3,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 102
},
{
"id": 392,
"affiliation": [
{
"affiliation": "Department of Emerging and Neglected Diseases, Biomedical Research Foundation, Dhaka, Bangladesh"
},
{
"affiliation": "Dhaka Medical College and Hospital, Dhaka, Bangladesh"
}
],
"first_name": "Nahida Hannan",
"family_name": "Nishat",
"email": null,
"author_order": 4,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 102
},
{
"id": 393,
"affiliation": [
{
"affiliation": "Department of Emerging and Neglected Diseases, Biomedical Research Foundation, Dhaka, Bangladesh"
},
{
"affiliation": "School of Environment and Life Science, Independent University Bangladesh (IUB), Dhaka, Bangladesh"
}
],
"first_name": "Mohammad Sorowar",
"family_name": "Hossain",
"email": "sorowar.hossain@brfbd.org",
"author_order": 5,
"ORCID": "https://orcid.org/0000-0001-7143-2909",
"corresponding": true,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "Mohammad Sorowar Hossain, PhD; Department of Emerging and Neglected Diseases, Biomedical Research Foundation, Dhaka, Bangladesh,\r\nEmail: sorowar.hossain@brfbd.org",
"article": 102
}
],
"views": 2996,
"downloads": 209,
"references": [
{
"id": 3152,
"serial_number": 1,
"pmc": null,
"reference": "Andersen KG, Rambaut A, Lipkin WI, Holmes EC, Garry RF. The proximal origin of SARS-CoV-2. Nat Med 2020;26:450 –2.",
"DOI": null,
"article": 102
},
{
"id": 8092,
"serial_number": 2,
"pmc": null,
"reference": "Lu R, Zhao X, Li J, Niu P, Yang B, Wu H, et al. Genomic characterisation and epidemiology of 2019 novel coronavirus: implications for virus origins and receptor binding. Lancet 2020;395:565–74.",
"DOI": null,
"article": 102
},
{
"id": 8093,
"serial_number": 3,
"pmc": null,
"reference": "Wiersinga WJ, Rhodes A, Cheng AC, Peacock SJ, Prescott HC. Pathophysiology, Transmission, Diagnosis, and Treatment of Coronavirus Disease 2019 (COVID-19): A Review. JAMA – J Am Med Assoc 2020;324:782–93.",
"DOI": null,
"article": 102
},
{
"id": 8094,
"serial_number": 4,
"pmc": null,
"reference": "Sudre CH, Lee K, Ni Lochlainn M, Varsavsky T, Murray B, Graham MS, et al. Symptom clusters in Covid19: A potential clinical prediction tool from the COVID Symptom study app. MedRxiv 2020:2020.06.12.20129056.",
"DOI": null,
"article": 102
},
{
"id": 8095,
"serial_number": 5,
"pmc": null,
"reference": "Chen J, Lau YF, Lamirande EW, Paddock CD, Bartlett JH, Zaki SR, et al. Cellular Immune Responses to Severe Acute Respiratory Syndrome Coronavirus (SARS-CoV) Infection in Senescent BALB/c Mice: CD4+ T Cells Are Important in Control of SARS-CoV Infection. J Virol 2010;84:1289–301.",
"DOI": null,
"article": 102
},
{
"id": 8096,
"serial_number": 6,
"pmc": null,
"reference": "COVID-19 Pandemic Planning Scenarios | CDC n.d. https://www.cdc.gov/coronavirus/2019-ncov/hcp/planning-scenarios.html (accessed August 4, 2020).",
"DOI": null,
"article": 102
},
{
"id": 8097,
"serial_number": 7,
"pmc": null,
"reference": "De Wit E, Van Doremalen N, Falzarano D, Munster VJ. SARS and MERS: Recent insights into emerging coronaviruses. Nat Rev Microbiol 2016;14:523–34.",
"DOI": null,
"article": 102
},
{
"id": 8098,
"serial_number": 8,
"pmc": null,
"reference": "Gupta A, Madhavan M V., Sehgal K, Nair N, Mahajan S, Sehrawat TS, et al. Extrapulmonary manifestations of COVID-19. Nat Med 2020;26.",
"DOI": null,
"article": 102
},
{
"id": 8099,
"serial_number": 9,
"pmc": null,
"reference": "Hamming I, Timens W, Bulthuis MLC, Lely AT, Navis GJ, van Goor H. Tissue distribution of ACE2 protein, the functional receptor for SARS coronavirus. A first step in understanding SARS pathogenesis. J Pathol 2004;203:631–7.",
"DOI": null,
"article": 102
},
{
"id": 8100,
"serial_number": 10,
"pmc": null,
"reference": "Ou X, Liu Y, Lei X, Li P, Mi D, Ren L, et al. Characterization of spike glycoprotein of SARS-CoV-2 on virus entry and its immune cross-reactivity with SARS-CoV. Nat Commun 2020;11:1–12.",
"DOI": null,
"article": 102
},
{
"id": 8101,
"serial_number": 11,
"pmc": null,
"reference": "hao Y, Zhao Z, Wang Y, Zhou Y, Ma Y, Zuo W. Single-cell RNA expression profiling of ACE2, the receptor of SARS-CoV-2. BioRxiv 2020:2020.01.26.919985.",
"DOI": null,
"article": 102
},
{
"id": 8102,
"serial_number": 12,
"pmc": null,
"reference": "Keidar S, Kaplan M, Gamliel-Lazarovich A. ACE2 of the heart: From angiotensin I to angiotensin (1-7). Cardiovasc Res 2007;73:463–9.",
"DOI": null,
"article": 102
},
{
"id": 8103,
"serial_number": 13,
"pmc": null,
"reference": "Wang K, Chen W, Zhou Y-S, Lian J-Q, Zhang Z, Du P, et al. SARS-CoV-2 invades host cells via a novel route: CD147-spike protein. BioRxiv 2020:2020.03.14.988345.",
"DOI": null,
"article": 102
},
{
"id": 8104,
"serial_number": 14,
"pmc": null,
"reference": "Hoffmann M, Kleine-Weber H, Schroeder S, Krüger N, Herrler T, Erichsen S, et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020;181:271-280.e8.",
"DOI": null,
"article": 102
},
{
"id": 8105,
"serial_number": 15,
"pmc": null,
"reference": "Chen YW, Yiu CPB, Wong KY. Prediction of the SARS-CoV-2 (2019-nCoV) 3C-like protease (3CLpro) structure: Virtual screening reveals velpatasvir, ledipasvir, and other drug repurposing candidates. F1000Research 2020;9.",
"DOI": null,
"article": 102
},
{
"id": 8106,
"serial_number": 16,
"pmc": null,
"reference": "Fehr AR, Perlman S. Coronaviruses: An overview of their replication and pathogenesis. Coronaviruses Methods Protoc., vol. 1282, Springer New York; 2015, p. 1–23.",
"DOI": null,
"article": 102
},
{
"id": 8107,
"serial_number": 17,
"pmc": null,
"reference": "Zhou P, Yang X Lou, Wang XG, Hu B, Zhang L, Zhang W, et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020;579:270–3.",
"DOI": null,
"article": 102
},
{
"id": 8108,
"serial_number": 18,
"pmc": null,
"reference": "Chau TN, Lee KC, Yao H, Tsang TY, Chow TC, Yeung YC, et al. SARS-Associated Viral Hepatitis Caused by a Novel Coronavirus: Report of Three Cases. Hepatology 2004;39:302–10.",
"DOI": null,
"article": 102
},
{
"id": 8109,
"serial_number": 19,
"pmc": null,
"reference": "Wölfel R, Corman VM, Guggemos W, Seilmaier M, Zange S, Müller MA, et al. Virological assessment of hospitalized patients with COVID-2019. Nature 2020;581:465–9.",
"DOI": null,
"article": 102
},
{
"id": 8110,
"serial_number": 20,
"pmc": null,
"reference": "Zhang H, Penninger JM, Li Y, Zhong N, Slutsky AS. Angiotensin-converting enzyme 2 (ACE2) as a SARS-CoV-2 receptor: molecular mechanisms and potential therapeutic target. Intensive Care Med 2020;46:586–90.",
"DOI": null,
"article": 102
},
{
"id": 8111,
"serial_number": 21,
"pmc": null,
"reference": "Imai Y, Kuba K, Rao S, Huan Y, Guo F, Guan B, et al. Angiotensin-converting enzyme 2 protects from severe acute lung failure. Nature 2005;436:112–6.",
"DOI": null,
"article": 102
},
{
"id": 8112,
"serial_number": 22,
"pmc": null,
"reference": "Marshall RP, Gohlke P, Chambers RC, Howell DC, Bottoms SE, Unger T, et al. Angiotensin II and the fibroproliferative response to acute lung injury. Am J Physiol – Lung Cell Mol Physiol 2004;286:156–64.",
"DOI": null,
"article": 102
},
{
"id": 8113,
"serial_number": 23,
"pmc": null,
"reference": "Tan WSD, Liao W, Zhou S, Mei D, Wong WSF. Targeting the renin–angiotensin system as novel therapeutic strategy for pulmonary diseases. Curr Opin Pharmacol 2018;40:9–17.",
"DOI": null,
"article": 102
},
{
"id": 8114,
"serial_number": 24,
"pmc": null,
"reference": "Kuba K, Imai Y, Rao S, Gao H, Guo F, Guan B, et al. A crucial role of angiotensin converting enzyme 2 (ACE2) in SARS coronavirus-induced lung injury. Nat Med 2005;11:875–9.",
"DOI": null,
"article": 102
},
{
"id": 8115,
"serial_number": 25,
"pmc": null,
"reference": "Ni W, Yang X, Yang D, Bao J, Li R, Xiao Y, et al. Role of angiotensin-converting enzyme 2 (ACE2) in COVID-19. Crit Care 2020;24:422.",
"DOI": null,
"article": 102
},
{
"id": 8116,
"serial_number": 26,
"pmc": null,
"reference": "Veerdonk F van de, Netea MG, Deuren M van, Meer JWM van der, Mast Q de, Bruggemann RJ, et al. Kinins and Cytokines in COVID-19: A Comprehensive Pathophysiological Approach. Preprints 2020:1–29.",
"DOI": null,
"article": 102
},
{
"id": 8117,
"serial_number": 27,
"pmc": null,
"reference": "Cohen AJ, DiFrancesco MF, Solomon SD, Vaduganathan M. Angioedema in COVID-19. Eur Heart J 2020.",
"DOI": null,
"article": 102
},
{
"id": 8118,
"serial_number": 28,
"pmc": null,
"reference": "Chen I-Y, Moriyama M, Chang M-F, Ichinohe T. Severe Acute Respiratory Syndrome Coronavirus Viroporin 3a Activates the NLRP3 Inflammasome. Front Microbiol 2019;10:50.",
"DOI": null,
"article": 102
},
{
"id": 8119,
"serial_number": 29,
"pmc": null,
"reference": "van den Berg DF, te Velde AA. Severe COVID-19: NLRP3 Inflammasome Dysregulated. Front Immunol 2020;11:1580.",
"DOI": null,
"article": 102
},
{
"id": 8120,
"serial_number": 30,
"pmc": null,
"reference": "Rothan HA, Byrareddy SN. The epidemiology and pathogenesis of coronavirus disease (COVID-19) outbreak. J Autoimmun 2020;109.",
"DOI": null,
"article": 102
},
{
"id": 8121,
"serial_number": 31,
"pmc": null,
"reference": "Coperchini F, Chiovato L, Croce L, Magri F, Rotondi M. The cytokine storm in COVID-19: An overview of the involvement of the chemokine/chemokine-receptor system. Cytokine Growth Factor Rev 2020;53:25–32.",
"DOI": null,
"article": 102
},
{
"id": 8122,
"serial_number": 32,
"pmc": null,
"reference": "Maurer M, Von Stebut E. Macrophage inflammatory protein-1. Int J Biochem Cell Biol 2004;36:1882–6.",
"DOI": null,
"article": 102
},
{
"id": 8123,
"serial_number": 33,
"pmc": null,
"reference": "azzucchelli RI, Warming S, Lawrence SM, Ishii M, Abshari M, Washington AV, et al. Visualization and Identification of IL-7 Producing Cells in Reporter Mice. PLoS One 2009;4:e7637.",
"DOI": null,
"article": 102
},
{
"id": 8124,
"serial_number": 34,
"pmc": null,
"reference": "Merad M, Martin JC. Pathological inflammation in patients with COVID-19: a key role for monocytes and macrophages. Nat Rev Immunol 2020;20:355–62.",
"DOI": null,
"article": 102
},
{
"id": 8125,
"serial_number": 35,
"pmc": null,
"reference": "Lotze MT, Tracey KJ. High-mobility group box 1 protein (HMGB1): Nuclear weapon in the immune arsenal. Nat Rev Immunol 2005;5:331–42.",
"DOI": null,
"article": 102
},
{
"id": 8126,
"serial_number": 36,
"pmc": null,
"reference": "Bardoel BW, Kenny EF, Sollberger G, Zychlinsky A. The balancing act of neutrophils. Cell Host Microbe 2014;15:526–36.",
"DOI": null,
"article": 102
},
{
"id": 8127,
"serial_number": 37,
"pmc": null,
"reference": "Zuo Y, Yalavarthi S, Shi H, Gockman K, Zuo M, Madison JA, et al. Neutrophil extracellular traps in COVID-19. JCI Insight 2020;5.",
"DOI": null,
"article": 102
},
{
"id": 8128,
"serial_number": 38,
"pmc": null,
"reference": "Zhou G, Chen S, Chen Z. Advances in COVID-19: the virus, the pathogenesis, and evidence-based control and therapeutic strategies. Front Med 2020;14:117–25.",
"DOI": null,
"article": 102
},
{
"id": 8129,
"serial_number": 39,
"pmc": null,
"reference": "Acharya D, Liu GQ, Gack MU. Dysregulation of type I interferon responses in COVID-19. Nat Rev Immunol 2020;20:397–8.",
"DOI": null,
"article": 102
},
{
"id": 8130,
"serial_number": 40,
"pmc": null,
"reference": "Channappanavar R, Fehr AR, Vijay R, Mack M, Zhao J, Meyerholz DK, et al. Dysregulated Type I Interferon and Inflammatory Monocyte-Macrophage Responses Cause Lethal Pneumonia in SARS-CoV-Infected Mice. Cell Host Microbe 2016;19:181–93.",
"DOI": null,
"article": 102
},
{
"id": 8131,
"serial_number": 41,
"pmc": null,
"reference": "Blanco-Melo D, Nilsson-Payant BE, Liu WC, Uhl S, Hoagland D, Møller R, et al. Imbalanced Host Response to SARS-CoV-2 Drives Development of COVID-19. Cell 2020;181:1036-1045.e9.",
"DOI": null,
"article": 102
},
{
"id": 8132,
"serial_number": 42,
"pmc": null,
"reference": "chen yongwen, Feng Z, Diao B, Wang R, Wang G, Wang C, et al. The Novel Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Directly Decimates Human Spleens and Lymph Nodes. MedRxiv 2020:2020.03.27.20045427.",
"DOI": null,
"article": 102
},
{
"id": 8133,
"serial_number": 43,
"pmc": null,
"reference": "Xu Z, Shi L, Wang Y, Zhang J, Huang L, Zhang C, et al. Pathological findings of COVID-19 associated with acute respiratory distress syndrome. Lancet Respir Med 2020;8:420–2.",
"DOI": null,
"article": 102
},
{
"id": 8134,
"serial_number": 44,
"pmc": null,
"reference": "Thevarajan I, Nguyen THO, Koutsakos M, Druce J, Caly L, van de Sandt CE, et al. Breadth of concomitant immune responses prior to patient recovery: a case report of non-severe COVID-19. Nat Med 2020;26:453–5.",
"DOI": null,
"article": 102
},
{
"id": 8135,
"serial_number": 45,
"pmc": null,
"reference": "Agrawal P, Nawadkar R, Ojha H, Kumar J, Sahu A. Complement evasion strategies of viruses: An overview. Front Microbiol 2017;8:1117.",
"DOI": null,
"article": 102
},
{
"id": 8136,
"serial_number": 46,
"pmc": null,
"reference": "Maglakelidze N, Manto KM, Craig TJ. A Review: Does Complement or the Contact System Have a Role in Protection or Pathogenesis of COVID-19? Pulm Ther 2020:1–8.",
"DOI": null,
"article": 102
},
{
"id": 8137,
"serial_number": 47,
"pmc": null,
"reference": "Abou-El-Hassan H, Zaraket H. Viral-derived complement inhibitors: current status and potential role in immunomodulation. Exp Biol Med 2017;242:397–410.",
"DOI": null,
"article": 102
},
{
"id": 8138,
"serial_number": 48,
"pmc": null,
"reference": "Carvelli J, Demaria O, Vély F, Batista L, Benmansour NC, Fares J, et al. Association of COVID-19 inflammation with activation of the C5a-C5aR1 axis. Nature 2020.",
"DOI": null,
"article": 102
},
{
"id": 8139,
"serial_number": 49,
"pmc": null,
"reference": "Magro C, Mulvey JJ, Berlin D, Nuovo G, Salvatore S, Harp J, et al. Complement associated microvascular injury and thrombosis in the pathogenesis of severe COVID-19 infection: a report of five cases. Transl Res 2020;220:1–13.",
"DOI": null,
"article": 102
},
{
"id": 8140,
"serial_number": 50,
"pmc": null,
"reference": "Edeas M, Saleh J, Peyssonnaux C. Iron: Innocent bystander or vicious culprit in COVID-19 pathogenesis? Int J Infect Dis 2020;97:303–5.",
"DOI": null,
"article": 102
},
{
"id": 8141,
"serial_number": 51,
"pmc": null,
"reference": "Phua J, Weng L, Ling L, Egi M, Lim CM, Divatia JV, et al. Intensive care management of coronavirus disease 2019 (COVID-19): challenges and recommendations. Lancet Respir Med 2020;8:506–17.",
"DOI": null,
"article": 102
},
{
"id": 8142,
"serial_number": 52,
"pmc": null,
"reference": "Mehta P, McAuley DF, Brown M, Sanchez E, Tattersall RS, Manson JJ. COVID-19: consider cytokine storm syndromes and immunosuppression. Lancet 2020;395:1033–4.",
"DOI": null,
"article": 102
},
{
"id": 8143,
"serial_number": 53,
"pmc": null,
"reference": "McDermid JM, Hennig BJ, van der Sande M, Hill AVS, Whittle HC, Jaye A, et al. Host iron redistribution as a risk factor for incident tuberculosis in HIV infection: An 11-year retrospective cohort study. BMC Infect Dis 2013;13:1–9.",
"DOI": null,
"article": 102
},
{
"id": 8144,
"serial_number": 54,
"pmc": null,
"reference": "Daher R, Manceau H, Karim Z. Iron metabolism and the role of the iron-regulating hormone hepcidin in health and disease. Press Medicale 2017;46:e272–8.",
"DOI": null,
"article": 102
},
{
"id": 8145,
"serial_number": 55,
"pmc": null,
"reference": "Kell DB, Pretorius E. Serum ferritin is an important inflammatory disease marker, as it is mainly a leakage product from damaged cells. Metallomics 2014;6:748–73.",
"DOI": null,
"article": 102
},
{
"id": 8146,
"serial_number": 56,
"pmc": null,
"reference": "Ursini F, Maiorino M. Lipid peroxidation and ferroptosis: The role of GSH and GPx4. Free Radic Biol Med 2020;152:175–85.",
"DOI": null,
"article": 102
},
{
"id": 8147,
"serial_number": 57,
"pmc": null,
"reference": "Yang WS, Stockwell BR. Ferroptosis: Death by Lipid Peroxidation. Trends Cell Biol 2016;26:165–76.",
"DOI": null,
"article": 102
},
{
"id": 8148,
"serial_number": 58,
"pmc": null,
"reference": "Polonikov A. Endogenous Deficiency of Glutathione as the Most Likely Cause of Serious Manifestations and Death in COVID-19 Patients. ACS Infect Dis 2020.",
"DOI": null,
"article": 102
},
{
"id": 8149,
"serial_number": 59,
"pmc": null,
"reference": "Parsanathan R, Jain SK. Glutathione deficiency induces epigenetic alterations of vitamin D metabolism genes in the livers of high-fat diet-fed obese mice. Sci Rep 2019;9:1–11.",
"DOI": null,
"article": 102
},
{
"id": 8150,
"serial_number": 60,
"pmc": null,
"reference": "Sun Y, Chen P, Zhai B, Zhang M, Xiang Y, Fang J, et al. The emerging role of ferroptosis in inflammation. Biomed Pharmacother 2020;127:110108.",
"DOI": null,
"article": 102
},
{
"id": 8151,
"serial_number": 61,
"pmc": null,
"reference": "Lamers MM, Beumer J, van der Vaart J, Knoops K, Puschhof J, Breugem TI, et al. SARS-CoV-2 productively infects human gut enterocytes. Science 2020;369:50–4.",
"DOI": null,
"article": 102
},
{
"id": 8152,
"serial_number": 62,
"pmc": null,
"reference": "Ahmed MU, Hanif M, Ali MJ, Haider MA, Kherani D, Memon GM, et al. Neurological Manifestations of COVID-19 (SARS-CoV-2): A Review. Front Neurol 2020;11:518.",
"DOI": null,
"article": 102
},
{
"id": 8153,
"serial_number": 63,
"pmc": null,
"reference": "Le Bon SD, Horoi M. Is anosmia the price to pay in an immune-induced scorched-earth policy against COVID-19? Med Hypotheses 2020;143:109881.",
"DOI": null,
"article": 102
},
{
"id": 8154,
"serial_number": 64,
"pmc": null,
"reference": "Oboti L, Peretto P, de Marchis S, Fasolo A. From chemical neuroanatomy to an understanding of the olfactory system. Eur J Histochem 2011;55:194–9.",
"DOI": null,
"article": 102
},
{
"id": 8155,
"serial_number": 65,
"pmc": null,
"reference": "Romero-Sánchez CM, Díaz-Maroto I, Fernández-Díaz E, Sánchez-Larsen Á, Layos-Romero A, García-García J, et al. Neurologic manifestations in hospitalized patients with COVID-19: The ALBACOVID registry. Neurology 2020:10.1212/WNL.0000000000009937.",
"DOI": null,
"article": 102
},
{
"id": 8156,
"serial_number": 66,
"pmc": null,
"reference": "Huang J, Zheng M, Tang X, Chen Y, Tong A, Zhou L. Potential of SARS-CoV-2 to Cause CNS Infection: Biologic Fundamental and Clinical Experience. Front Neurol 2020;11:659.",
"DOI": null,
"article": 102
},
{
"id": 8157,
"serial_number": 67,
"pmc": null,
"reference": "Gholizadeh P, Safari R, Marofi P, Zeinalzadeh E, Pagliano P, Ganbarov K, et al. Alteration of Liver Biomarkers in Patients with SARS-CoV-2 (COVID-19). J Inflamm Res 2020;Volume 13:285–92.",
"DOI": null,
"article": 102
},
{
"id": 8158,
"serial_number": 68,
"pmc": null,
"reference": "Hirsch JS, Ng JH, Ross DW, Sharma P, Shah HH, Barnett RL, et al. Acute kidney injury in patients hospitalized with COVID-19. Kidney Int 2020;98:209–18.",
"DOI": null,
"article": 102
},
{
"id": 8159,
"serial_number": 69,
"pmc": null,
"reference": "Tisoncik JR, Korth MJ, Simmons CP, Farrar J, Martin TR, Katze MG. Into the Eye of the Cytokine Storm. Microbiol Mol Biol Rev 2012;76:16–32.",
"DOI": null,
"article": 102
},
{
"id": 8160,
"serial_number": 70,
"pmc": null,
"reference": "Zhang B, Zhou X, Qiu Y, Feng F, Feng J, Jia Y, et al. Clinical characteristics of 82 death cases with COVID-19. MedRxiv 2020:2020.02.26.20028191.",
"DOI": null,
"article": 102
},
{
"id": 8161,
"serial_number": 71,
"pmc": null,
"reference": "Yuki K, Fujiogi M, Koutsogiannaki S. COVID-19 pathophysiology: A review. Clin Immunol 2020;215:108427.",
"DOI": null,
"article": 102
},
{
"id": 8162,
"serial_number": 72,
"pmc": null,
"reference": "Lucas C, Wong P, Klein J, Castro TBR, Silva J, Sundaram M, et al. Longitudinal analyses reveal immunological misfiring in severe COVID-19. Nature 2020:1–9.",
"DOI": null,
"article": 102
},
{
"id": 8163,
"serial_number": 73,
"pmc": null,
"reference": "Bourgonje AR, Abdulle AE, Timens W, Hillebrands J, Navis GJ, Gordijn SJ, et al. Angiotensin‐converting enzyme 2 (ACE2), SARS‐CoV‐2 and the pathophysiology of coronavirus disease 2019 (COVID‐19). J Pathol 2020;251:228–48.",
"DOI": null,
"article": 102
},
{
"id": 8164,
"serial_number": 74,
"pmc": null,
"reference": "Goshua G, Pine AB, Meizlish ML, Chang CH, Zhang H, Bahel P, et al. Endotheliopathy in COVID-19-associated coagulopathy: evidence from a single-centre, cross-sectional study. Lancet Haematol 2020;7:e575–82.",
"DOI": null,
"article": 102
},
{
"id": 8165,
"serial_number": 75,
"pmc": null,
"reference": "Gupta N, Zhao YY, Evans CE. The stimulation of thrombosis by hypoxia. Thromb Res 2019;181:77–83.",
"DOI": null,
"article": 102
},
{
"id": 8166,
"serial_number": 76,
"pmc": null,
"reference": "Bompard F, Monnier H, Saab I, Tordjman M, Abdoul H, Fournier L, et al. Pulmonary embolism in patients with Covid-19 pneumonia. Eur Respir J 2020:2001365.",
"DOI": null,
"article": 102
},
{
"id": 8167,
"serial_number": 77,
"pmc": null,
"reference": "Wu C, Chen X, Cai Y, Xia J, Zhou X, Xu S, et al. Risk Factors Associated With Acute Respiratory Distress Syndrome and Death in Patients With Coronavirus Disease 2019 Pneumonia in Wuhan, China. JAMA Intern Med 2020;180:934.",
"DOI": null,
"article": 102
},
{
"id": 8168,
"serial_number": 78,
"pmc": null,
"reference": "Zhou F, Yu T, Du R, Fan G, Liu Y, Liu Z, et al. Clinical course and risk factors for mortality of adult inpatients with COVID-19 in Wuhan, China: a retrospective cohort study. Lancet 2020;395:1054–62.",
"DOI": null,
"article": 102
},
{
"id": 8169,
"serial_number": 79,
"pmc": null,
"reference": "Jiang L, Tang K, Levin M, Irfan O, Morris SK, Wilson K, et al. COVID-19 and multisystem inflammatory syndrome in children and adolescents. Lancet Infect Dis 2020;0.",
"DOI": null,
"article": 102
}
]
},
{
"id": 107,
"slug": "178-1597540568-qualitative-phytochemicals-and-pharmacological-properties-analysis-of-methanol-extract-of-tabernaemontana-pauciflora-leaves",
"featured": false,
"slider": false,
"issue": "Vol4 Issue1",
"type": "original_article",
"manuscript_id": "178-1597540568",
"recieved": "2020-07-21",
"revised": null,
"accepted": "2020-08-31",
"published": "2020-09-15",
"pdf_file": "https://jabet.bsmiab.org/media/pdf_file/2023/39/178-1597540568.pdf",
"title": "Qualitative phytochemicals and pharmacological properties analysis of methanol extract of Tabernaemontana pauciflora leaves",
"abstract": "<p>The methanol extract of the leaves of <em>Tabernaemontana pauciflor</em>a (METP) was screened to investigate the qualitative phytochemical, <em>in vivo</em> anti-depressant, anxiolytic and <em>in vitro</em> thrombolytic activities. The <em>in vivo</em> antidepressant activity of METP was examined by force swimming test (FST) and tail suspension test (TST), whereas anxiolytic activity was examined by using hole-board test (HBT) and elevated plus maze test (EPM) on mice model. Additionally, qualitative phytochemical assay was performed to identify some secondary metabolites, namely alkaloids, carbohydrates, flavonoids. The <em>in vitro</em> thrombolytic model was used to evaluate the clot lysis effect of METP. In the antidepressant assay, METP (200 and 400 mg/kg) exhibited significant anti-depression activity <em>(P < 0.001</em>) on FST (45.64% and 58.31%) and TST (38.67% and 63.62%) whereas the notable anxiolytic effect was observed in EPM (67.67% and 56.06%) and HBT (35.53% and 50%) in dose dependent manner which compared with control. In <em>in vitro</em> thrombolytic assessment of METP exposed 22.66% inhibition of clot. The current results proposed that METP might be effective in neuropharmacological and thrombolytic response due to the presence of alkaloids, tannins and flavonoids, which required further studies to investigate the mechanism.</p>",
"journal_reference": "J Adv Biotechnol Exp Ther. 2021; 4(1): 09-16.",
"academic_editor": "Akhi Moni, PhD; ABEx Bio-Research Center, Bangladesh",
"cite_info": "Hossen A, Arman M, et al. Qualitative phytochemicals and pharmacological properties analysis of methanol extract of Tabernaemontana pauciflora leaves. J Adv Biotechnol Exp Ther. 2021; 4(1): 09-16.",
"keywords": [
"Tabernaemontana pauciflora",
"Anti-depressant",
"Thrombolytic",
"Anxiolytic"
],
"DOI": "10.5455/jabet.2021.d101",
"sections": [
{
"section_number": 1,
"section_title": "INTRODUCTION",
"body": "<p>Neurological disorders are the common phenomena. Currently an estimated 14.7M neurological cases, which according to 2019 is equivalent to at least 1 in 6 individuals living with one or more sign of neurological problems [<a href=\"#r-1\">1</a>, <a href=\"#r-2\">2</a>]. Benzodiazepines are synthetic drugs to prescribe frequently for their anxiolytic, sedative-hypnotic and anticonvulsant actions [<a href=\"#r-3\">3</a>]. Besides, depression is a common and serious dominant mental disorder that negatively affects how you feel on character, apathy, loss of energy, interruption of thinking and activity, as well as profound feelings of frustration, despair and suicidal ideation [<a href=\"#r-3\">3</a>]. Instead of the availability of antidepressant drugs like TCA (Tricyclic antidepressant), selective reversible inhibitors of monoamine oxidase-A (MAO-A), selective serotonin reuptake inhibitors (SSRIs) and selective noradrenalin reuptake inhibitors (SNRIs) [<a href=\"#r-4\">4</a>]. Depression is still denoted as a major medical problem associated with major side effects [<a href=\"#r-5\">5</a>]. Not only associated with physical dependence and tolerance providing side effects, these psycho-neural drugs also cause for harmful effects on respiratory system, digestive system and immune systems of the human organ [<a href=\"#r-6\">6</a>].<br />\r\nThe depression is often conducted by irritability, anxiety, confusion, insomnia and behavioral disorders. As a result, the sequential use of an anxiolytic with an antidepressant agent is often necessary to recover the disease [<a href=\"#r-7\">7</a>]. Selective serotonin reuptake inhibitors (SSRIs) are currently used as the first-choice therapy for depression, but insomnia, irritability and anxiety sometimes occur in the early stages of treatment. As these symptoms may result in early dropout or delays in the anti-depressive response, the concomitant use of an anxiolytic is considered helpful. A new anxiolytic and antidepressant agents that are why to search with the lowest adverse effect is still researchers have high interest of this area [<a href=\"#r-8\">8</a>]<br />\r\nAtherothrombotic, the formation of a blood clot within an artery as a result of atherosclerosis named cardiovascular disease are characterized by the existence of great influences of thrombus in the blood vessel [<a href=\"#r-9\">9</a>]. The formation of complex form by connecting among platelets to platelets, also to leucocytes by activated platelets results in plaque formation and development of thrombosis. A neutral agent called plasmin triggers clot lysis by crumble up fibrinogen & fibrin contained clot [<a href=\"#r-10\">10</a>]. Additional plasminogen agent can easily convert to plasmin by streptokinase enzyme which forms a 1:1 stoichiometric complex with plasminogen [<a href=\"#r-11\">11</a>].<br />\r\n<em>T. pauciflora </em>of the family of Apocynaceae locally named as Bunut, Malul-ulat, Rodok. It is distributed in Chittagong hill tracts area, Bangladesh, Vietnam, Cambodia, peninsular Malaysia (widespread), Myanmar, Borneo. Traditionally the different part of <em>T. pauciflora</em> is used to cure headaches, to remedy various syphilitic afflictions effect and to treat eruptions of the skin [<a href=\"#r-12\">12</a>]. Based on their ethno botanical uses the present study aimed to investigate the qualitative phytochemical analysis, anxiolytic, antidepressant and thrombolytic activities of methanol extract of <em>T. Pauciflora </em>leaves.</p>"
},
{
"section_number": 2,
"section_title": "MATERIALS AND METHODS",
"body": "<p><strong>Chemicals</strong><br />\r\nThe chemicals were used for the experiments as methanol (Sigma Chemicals, USA). Diazepam, fluoxetine (Eskayef Banlgadesh Ltd; Tongi, Bangladesh) and lyophilized streptokinase (SK) vial (Square Pharmaceuticals Ltd. Bangladesh). All other chemicals were obtained from local trader through Taj Scientific Ltd, Chittagong, Bangladesh.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Animals</strong><br />\r\nThe study was accompanied on Swiss Albino mice were purchased from Jahangirnagar University, Dhaka, Bangladesh. They were five to six weeks of age of male-female, weighing about 20 – 30 g, which were kept in housed colony cages (six mice per cages) at ambient temperature of 25-27°C with 12 hours light and dark cycles having with proper ventilation in the room [<a href=\"#r-13\">13</a>]. The study was conducted according to the guidelines of P&D Committee of Department of Pharmacy, International Islamic University Chittagong (Pharm/AEC/138/13-’19, 22/12/2019).</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Collection and preparation of plant extract</strong><br />\r\nFresh leaves of <em>T. pauciflora</em> were collected from a local village Chittagong, Bangladesh and authenticated by Dr Shaikh Bokhtiar Uddin, Professor, Department of Botany, University of Chittagong. The leaves were shaded dried for a multi-week time and ground into a fine powder utilizing a process machine. The ground leaves (1000g) were absorbed adequate measure of methanol (3L) for ten days at room temperature with incidental shaking and mixing at that point separated through with cotton plug followed by Whitman channel paper No. 1. After filtration, the filtrated was subjected for the evaporation in water bath. The obtained semisolid yield extract was 10.06% which was refrigerated at 4 degree centigrade for further use.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Qualitative phytochemical analysis</strong><br />\r\nThe METP tried for the screening of bioactive mixes by subjective way utilizing keeping standard method to assess the alkaloids, carbohydrates, flavonoids, terpenoids, tannins, polyphenol, phenols, steroids, amides and glycosides [<a href=\"#r-13\">13-15</a>].</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Evaluation of anxiolytic activity</strong><br />\r\n<em>Elevated plus maze (EPM) test</em><br />\r\nThe EPM test is used mainly to assess exploratory behaviors in a mouse model. The EPM apparatus consisted of two open arms (5×10 cm<sup>2</sup>) separated by two closed arms (5×10×15 cm<sup>3</sup>), which raised to a height of 25 cm in a dimly light space [<a href=\"#r-16\">16</a>, <a href=\"#r-17\">17</a>]. The maze floor and walls were constructed from dark opaque wood. Mice (n = 5) were treated with extract (200 and 400 mg/kg). After 30 min, each mouse placed individually in the center of the EPM facing towards closed arm and recorded the % of time spent in the open arms for 5 min.<br />\r\n% time spent in open arm = (time spent in open arm)/(time spent in open arm + time spent in closed arm) ×100</p>\r\n\r\n<p> </p>\r\n\r\n<p><em>Hole-board test (HBT)</em><br />\r\nThe hole-board apparatus was applied for this experiment as the method stated earlier. The tool consisted of a wooden box (40 cm × 40 cm.) in diameter with 16 holes uniformly distributed on the surface. The apparatus elevated to the 25 cm height. In brief, mice were tested after the 30 minutes of dosing with METP (200 and 400 mg/kg) and 15 minutes after the administration of diazepam (1 mg/kg), whereas each mice was allowed to move on the platform and the number of head dips in the holes was counted for 5 minutes [<a href=\"#r-18\">18</a>, <a href=\"#r-19\">19</a>].</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Evaluation of anti-depressant activity</strong><br />\r\n<em>Force swimming test (FST)</em><br />\r\nForce swimming test is the most widely used assay described by Porsolt <em>et. al</em>. frequently used behavioral models for screening antidepressant activity. The animals were divided into four groups with 5 in each and prepared the test dose 400 mg/kg and 200 mg/kg along with control dose (0.9% saline water; 10 ml/ kg orally) and fluoxetine hydrochloride (10 mg/kg). Then take the mice’s weight and administrate dose according to their weight. After 30 minutes each mouse was individually forced to swim in open glass chamber (25 × 15 × 25 cm) containing fresh water to a height of 15 cm and maintained at 26° ± 1°C. It has to be ensured that mice could not get any support by touching the bottom of the sidewalls of the chamber. Each of the mice shows vigorous movement during the initial 2 minutes period of the test. The duration of immobility was manually recorded during the next 5 minutes of the total 7 min testing period [<a href=\"#r-20\">20</a>, <a href=\"#r-21\">21</a>].</p>\r\n\r\n<p> </p>\r\n\r\n<p><em>Tail suspension test (TST)</em><br />\r\nThis procedure test commonly employed behavioral model for screening antidepressant like activity in mice was first given by Stern <em>et al.</em> The animals are divided into four groups with 5 in each. At first, we have to prepare the test dose 400 mg/kg and 200 mg/kg along with control dose (0.9% saline water, 10 ml/kg orally) and Fluoxetine hydrochloride (10 mg/kg). Then take the mice’s weight and administrate dose according to their weight. After 30 minutes of dose administration individually each mouse was suspended to the edge of a table, the table was 50 cm above the floor and tape was placed approximately 1 cm from the tip of the tail. The total period of immobility was recorded manually for 7 min [<a href=\"#r-22\">22</a>, <a href=\"#r-23\">23</a>].</p>\r\n\r\n<p> </p>\r\n\r\n<p><em>Evaluation of thrombolytic activity</em><br />\r\nThe thrombolytic effect of METP justified through slide modification of method which described by Daginawala and Kawsar <em>et al</em>. [<a href=\"#r-24\">24</a>, <a href=\"#r-25\">25</a>], where streptokinase was used as standard. Streptokinase vial (15, 00,000 I.U) which are available in drug market was accepted and prepared solution with 5 ml distilled water. Later, streptokinase 100 μl dose from this stock solution were used for in vitro analysis. Blood was derived from five healthy students except consideration of previous antithrombotic therapy. After that, collected blood was assigned into pre-weighed micro centrifuge tube for allowing clot formation. The following formula was applied for the determination of clot lysis:<br />\r\nClot lysis (%) = (weight of released clot/clot weight) × 100</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Statistical analysis</strong><br />\r\nThe values were showed in mean ± standard error mean (SEM). <em>**P< 0.001 </em>and<em> *P< 0.01</em> statistically significant, which was carried by one-way ANOVA (Dunnett’s test) using GraphPad Prism (version 8.4.) software.</p>"
},
{
"section_number": 3,
"section_title": "RESULTS",
"body": "<p><strong>Qualitative phytochemical assay</strong><br />\r\nPhytochemicals are present and the total content of METP is alkaloids, carbohydrates, tannins, flavonoids, steroids and amides presented in the <a href=\"#Table-1\">Table 1</a>. It was found to be absent of glycosides, polyphenol, phenols and terpenoids in the METP.</p>\r\n\r\n<div id=\"Table-1\">\r\n<p><a href=\"https://jabet.bsmiab.org/table/178-1597540568-table1/\">Table-1</a><strong>Table 1. </strong>Qualitative phytochemical screening of <em>T. pauciflora</em> (leaves).</p>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Effect of methanol extract of T. pauciflora on anxiolytic activity</strong><br />\r\nThe METP displayed increased percentage of time spent into open arms, exposed in <a href=\"#figure1\">Figure 1 (A)</a>. The METP displayed a significant percentage of time spent in open arm (67.67 ± 1.45 and 56.06 ± 2.44) by 400 and 200 mg/kg respectively compared with control (55.06 ± 2.77), whereas the standard drug diazepam exhibited (87.02 ± 2.08). Additionally, HBT is an exploratory technique applied in animal to measure anxiety, stress and emotion. There was dose depended effect increases when mice treated with METP 400 mg/kg and 200 mg/kg, represented in <a href=\"#figure1\">Figure 1 (B)</a>. The METP 400 mg/kg dose exhibited significant number of head dipping (50 ± 2.65) compared to control (25.67± 2.33), whereas the diazepam showed (66 ± 1.73) number of head dipping in hole-board apparatus.</p>\r\n\r\n<div id=\"figure1\">\r\n<figure class=\"image\"><img alt=\"\" height=\"201\" src=\"/media/article_images/2024/21/03/178-1597540568-Figure1.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 1. </strong>(A) Anxiolytic action of methanol extract of T. pauciflora (leaves), control and diazepam on percentage of time spent in open arm in elevated plus maze test (EPM). (B) Anxiolytic effect of methanol extract of T. pauciflora (leaves), diazepam and control on number of head-dipping in hole-board test (HBT). Values are represented in Mean ± SEM (n = 5). In both test METP dose 400 mg/kg exhibited P values (**P< 0.001) is statistically significant in comparison to control followed by Dunnett test of one-way ANOVA. METP = Methanol extract of <em>Tabernaemontana pauciflora</em> (leaves).</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Effect of methanol extract of </strong><strong><em>T. pauciflora</em></strong><strong> on antidepressant activity</strong><br />\r\nPossible antidepressant effect of METP was tested in the FST after oral administration. In this study, animals treated with two doses of METP (200 and 400 mg/kg) reported decreases in their time of immobility, which was important for METP at (49.33 ± 2.60 s and 64.33 ± 2.03 s), compared to control (118.33 ± 2.60 s and 22.67 ± 1.45 s). The result presented in <a href=\"#figure2\">Figure 2 (A)</a>. Similarly, as predicted, animals were treated with fluoxetine (10 mg/kg) showed a substantial decrease (22.6 s) in the immobility period. The mice was treated with two doses of METP 200 and 400 mg/kg in this FST, revealed decreases in their immobility times, which was (98.33 ± 0.88 s and 56 ± 0.88 s) for methanol extract of <em>T. pauciflora</em> (leaves) when compared to negative control (160.33 ± 1.45 s), shown in <a href=\"#figure2\">Figure 2 (B)</a>. Similarly, mice treated with standard fluoxetine (10 mg/kg) as expected, manifested a remarkable decrease in the immobility time (20.66 ± 1.45 s).</p>\r\n\r\n<div id=\"figure2\">\r\n<figure class=\"image\"><img alt=\"\" height=\"182\" src=\"/media/article_images/2024/21/03/178-1597540568-Figure2.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 2. </strong>(A) Antidepressant activity of methanol extract of <em>T. pauciflora</em> (leaves) and fluoxetine on force swimming test (FST). (B) Antidepressant effect of methanol extract of <em>T. pauciflora</em> (leaves) and fluoxetine on tail suspension test (TST). Values are represented in Mean ± SEM (n = 5). <sup>**</sup><em>P</em>< 0.001 statistically significant in comparison to control followed by Dunnett test of one-way ANOVA. METP = Methanol extract of <em>Tabernaemontana pauciflora</em> (leaves).</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Effect of methanol extract of </strong><strong><em>T. pauciflora</em></strong><strong> on thrombolytic activity</strong><br />\r\nThe <em>in vitro</em> thrombolytic model was used to measure the clot lysis of the METP compared with streptokinase, and normal saline (control) expressed in <a href=\"#figure3\">Figure 3</a>. The METP exposed significant (<em>P </em>< 0.001) percentage of clot lysis activity (22.66 ± 1.96%) compared to control (4.66 ± 0.63%), whereas standard drug streptokinase evinced (75.0 ± 1.14%) clot lysis action.</p>\r\n\r\n<div id=\"figure3\">\r\n<figure class=\"image\"><img alt=\"\" height=\"329\" src=\"/media/article_images/2024/26/03/178-1597540568-Figure3.jpg\" width=\"442\" />\r\n<figcaption><strong>Figure 3. </strong>The clot lysis activity of methanol extract of <em>T. pauciflora</em> (leaves), streptokinase and normal saline (Control). Values are represented in Mean ± SEM (n = 5). METP dose displayed P value (<sup>**</sup><em>P</em>< 0.001) is statistically significant in comparison to control followed by Dunnett test of one-way ANOVA. METP = Methanol extract of <em>Tabernaemontana pauciflora</em> (leaves).</figcaption>\r\n</figure>\r\n</div>"
},
{
"section_number": 4,
"section_title": "DISCUSSION",
"body": "<p>The drug discovery system is the process in which new candidate medications are discovered. Medicinal plant drug discovery continuously increasing day by day to provide new and significant leads against pharmacological targets [<a href=\"#r-26\">26</a>]. Phytochemical investigation of the crude extract was done to find out the uncovered presence of compound which might be responsible for the therapeutic uses [<a href=\"#r-27\">27</a>, <a href=\"#r-28\">28</a>]. The phytochemical (qualitative) studies demonstrated that the methanolic extracts of <em>T. pauciflora </em>contained alkaloids, tannins, flavonoids, steroids, and amides. This phytochemical group present in the METP may help us to find out the specific compound and also help us to estimate how much bioactive in this plant extract.<br />\r\nDepressant causes a slowing motion in the physiological functions. There is various leading cause falling in depression such as oxidative stress, long time inflammation, and several chronic disorders, trauma hemorrhage, difficulties within the family, which causes a slowing motion in the physiological functions [<a href=\"#r-29\">29</a>, <a href=\"#r-30\">30</a>]. Antidepressants are those medications that help relieve symptoms of mood disorders by repairing neurotransmitter chemical imbalances in the brain. The tail suspension test (TST) is done to predict the antidepressant pharmacological activity in animal model [<a href=\"#r-31\">31</a>]. The total immobility time after induced by tail suspension of METP was measured according by the Steru <em>et al.</em> [<a href=\"#r-31\">31</a>, <a href=\"#r-32\">32</a>]. In this test animal were treated with two doses of METP 200 and 400 mg/kg, results are represented as mean ± SEM, where the METP 400 mg/kg exhibited significant decreases in the immobility times as (58.33 ± 1.76 s) compared with the control as (160.33 ± 1.45 s), whereas the standard drug fluoxetin showed (20.33 ± 1.76 s) immobility time [<a href=\"#r-33\">33</a>]. Force swimming test is commonly used to screen antidepressant activity and other types of psychotropic actions in animal [<a href=\"#r-34\">34</a>]. The term immobility are considered to possess the behavioral desperation analogous to that seen in human depression and it is well endowed that antidepressant ligands are able to inhibit the duration of immobility in animal [<a href=\"#r-35\">35</a>]. The both dose of METP displayed remarkable reduction of immobility time (49.33 ± 2.60 s and 64.33 ± 2.03 s) compared with the control (118.33 ± 2.60 s). The METP 400 mg/kg displayed significant decreases of number of head dipping (50 ± 2.65) compared with control (25.67± 2.33) 10 ml/kg whereas standard (Diazepam) 1 mg/kg showed (66 ± 1.73). From above discussion it can be suggested that the methanol extract of <em>T. pauciflora</em> (precisely METP 400 mg/kg) has significant anti-depressant effect.<br />\r\nAnxiety denotes a normal reaction to stress. Anxiolytics relieve anxiety disorders associated with rapid heart rate, palpitations and chest pain and also can increase the risk of high blood pressure and cardiac disease [<a href=\"#r-36\">36</a>, <a href=\"#r-37\">37</a>]. The Elevated Plus Maze (EPM) test has been used in a model of anxiety [<a href=\"#r-38\">38</a>]. In the EPM test, the METP 400 mg/kg dose revealed crucial inhibition of number of time spent (67.67 ± 1.45) compared to control (1% tween-80 solution) dose. In this test diazepam is the reference control or positive control with anxiolytic effect mainly used in the chronic treatment [<a href=\"#r-39\">39</a>, <a href=\"#r-40\">40</a>]. The HBT is an experimental system which was designed 1970s as open arm test and done in animal to evaluate anxiolytic result [<a href=\"#r-41\">41</a>] by measuring anxiety and emotion. In this study , we use this hole board test method to assess the new leads for anxiolytic activity compared to standard (reference) drug diazepam in animal model [<a href=\"#r-42\">42</a>]. Every mouse evaluate by placing it separately in the center of the hole board apparatus and observed its activity for head dipping of the mouse is determined when both ears below the top of the hole [<a href=\"#r-43\">43</a>]. The extract dose has anxiolytic activity determined by the reduction of number of head dipping with time spent after exposer of the METP dose compared with the control [<a href=\"#r-44\">44</a>, <a href=\"#r-45\">45</a>]. The both METP 400 mg/kg and 200 mg/kg displayed significant decreases of number of head dipping (84 ± 2.65 and 100 ± 3.21) compared with control (1% tween-80 solution) 10 ml/kg whereas standard (Diazepam) 1 mg/kg showed (13 ± 1.73). From above discussion, we may can displayed another types anxiolytic ligand which will be effective if this result may analysis further. For the evaluation of leads compound of anxiolytic activity test method the elevated plus maze test and hole-board test were used. The presence of anxiolytic effect of METP due to the presence of flavonoid group [<a href=\"#r-46\">46</a>].<br />\r\nThrombolysis is a parameter which dissolve the blood clot already fromed in the blood vessel and protect from the various heart diseases through improving blood flow, especially infused an artificial enzyme such as streptokinase in dangerous situation associated with major pulmonary embolism, late arterial occlusion and in deep vein thrombosis [<a href=\"#r-47\">47</a>, <a href=\"#r-48\">48</a>]. Activation of blood lysis factors such as plasmin,fibrinogen and fibrin and others enhances the breakdown of the blood clot to increase the blood flow [<a href=\"#r-49\">49</a>, <a href=\"#r-50\">50</a>]. Generally, thrombolytic agent activates the plasminogen that forms plasmin which acts as a cleaved product to dissolve blood clots [<a href=\"#r-51\">51</a>]. In this project we studied <em>in vitro</em> trombolytic experiment of METP, where METP exhibited siginifant percent of clot lysis (22.67 ± 1.96) applied in animal model compared with the control (4.66 ± 0.63), whereas standard drug manifested effective result <em>(**P</em>< 0.001). This effect may showed due to presence of flavonoids contents in the extract [<a href=\"#r-52\">52</a>]. This significant result compared to negative control due to the presence of alkaloids and tannins [<a href=\"#r-53\">53</a>].</p>\r\n\r\n<div id=\"figure4\">\r\n<figure class=\"image\"><img alt=\"\" height=\"331\" src=\"/media/article_images/2024/21/03/178-1597540568-Figure4.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 4.</strong> Graphical representation of methanol extract of T. pauciflora (leaves) on qualitative phytochemical, antidepressant, anxiolytic and thrombolytic activity.</figcaption>\r\n</figure>\r\n</div>"
},
{
"section_number": 5,
"section_title": "CONCLUSION",
"body": "<p>In our present well-informed study stated that methanol extract of <em>T. pauciflora</em> is potetial source for the important phytochemicals having anxiolytic, antidepressant and thrombolytic outcome (<a href=\"#figure4\">Figure 4</a>). According to study of the results and discussions of METP, this plant may contain effective herbal drug. Further advanced studies are required to carefully evaluate the mechanisms which responsible for this anxiolytic and antidepressants phenomenon being reported.</p>"
},
{
"section_number": 6,
"section_title": "ACKNOWLEDGEMENTS",
"body": "<p>Authors are thankful to Department of Pharmacy, International Islamic University Chittagong, Chittagong, Bangladesh, for their research facilities and support. The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request. This work is conducted with the individual funding of all authors.</p>"
},
{
"section_number": 7,
"section_title": "CONFLICTS OF INTERESTS",
"body": "<p>We, the authors, declare that we have no competing interests.</p>"
},
{
"section_number": 8,
"section_title": "AUTHOR CONTRIBUTIONS",
"body": "<p>AH, MNI and AU together planned and designed the research. MNI arranged the whole facilities and co-supervised this research. AH conducted the entire laboratory works. MA, NB, NA, SAS and AH imparted in study design and interpreted the results putting efforts on statistical analysis with MRAM, SAJS and MNI participated in the manuscript draft and has thoroughly checked and revised the manuscript for necessary changes in format, grammar and English standard. All authors read and agreed on the final version of the manuscript.</p>"
}
],
"figures": [
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/21/03/178-1597540568-Figure1.jpg",
"caption": "Figure 1. (A) Anxiolytic action of methanol extract of T. pauciflora (leaves), control and diazepam on percentage of time spent in open arm in elevated plus maze test (EPM). (B) Anxiolytic effect of methanol extract of T. pauciflora (leaves), diazepam and control on number of head-dipping in hole-board test (HBT). Values are represented in Mean ± SEM (n = 5). In both test METP dose 400 mg/kg exhibited P values (**P< 0.001) is statistically significant in comparison to control followed by Dunnett test of one-way ANOVA. METP = Methanol extract of Tabernaemontana pauciflora (leaves).",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/21/03/178-1597540568-Figure2.jpg",
"caption": "Figure 2. (A) Antidepressant activity of methanol extract of T. pauciflora (leaves) and fluoxetine on force swimming test (FST). (B) Antidepressant effect of methanol extract of T. pauciflora (leaves) and fluoxetine on tail suspension test (TST). Values are represented in Mean ± SEM (n = 5). **P< 0.001 statistically significant in comparison to control followed by Dunnett test of one-way ANOVA. METP = Methanol extract of Tabernaemontana pauciflora (leaves).",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/26/03/178-1597540568-Figure3.jpg",
"caption": "Figure 3. The clot lysis activity of methanol extract of T. pauciflora (leaves), streptokinase and normal saline (Control). Values are represented in Mean ± SEM (n = 5). METP dose displayed P value (**P< 0.001) is statistically significant in comparison to control followed by Dunnett test of one-way ANOVA. METP = Methanol extract of Tabernaemontana pauciflora (leaves).",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/21/03/178-1597540568-Figure4.jpg",
"caption": "Figure 4. Graphical representation of methanol extract of T. pauciflora (leaves) on qualitative phytochemical, antidepressant, anxiolytic and thrombolytic activity.",
"featured": false
}
],
"authors": [
{
"id": 415,
"affiliation": [
{
"affiliation": "Department of Pharmacy, International Islamic University Chittagong, Chittagong-4318, Bangladesh"
}
],
"first_name": "Anowar",
"family_name": "Hossen",
"email": null,
"author_order": 1,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 107
},
{
"id": 416,
"affiliation": [
{
"affiliation": "Department of Pharmacy, International Islamic University Chittagong, Chittagong-4318, Bangladesh"
}
],
"first_name": "Mohammad",
"family_name": "Arman",
"email": null,
"author_order": 2,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 107
},
{
"id": 417,
"affiliation": [
{
"affiliation": "Department of Pharmacy, International Islamic University Chittagong, Chittagong-4318, Bangladesh"
}
],
"first_name": "Syed Al Jawad",
"family_name": "Sayem",
"email": null,
"author_order": 3,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 107
},
{
"id": 418,
"affiliation": [
{
"affiliation": "Department of Pharmacy, International Islamic University Chittagong, Chittagong-4318, Bangladesh"
}
],
"first_name": "Mohammad Rifat Alam",
"family_name": "Maruf",
"email": null,
"author_order": 4,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 107
},
{
"id": 419,
"affiliation": [
{
"affiliation": "Department of Pharmacy, International Islamic University Chittagong, Chittagong-4318, Bangladesh"
}
],
"first_name": "Md. Ahsan",
"family_name": "Ullah",
"email": null,
"author_order": 5,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 107
},
{
"id": 420,
"affiliation": [
{
"affiliation": "Department of Pharmacy, International Islamic University Chittagong, Chittagong-4318, Bangladesh"
}
],
"first_name": "Mohammad Nazmul",
"family_name": "Islam",
"email": "sayeadiiuc@gmail.com",
"author_order": 6,
"ORCID": null,
"corresponding": true,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "Mohammad Nazmul Islam, Department of Pharmacy, International Islamic University Chittagong, Chittagong-4318, Bangladesh, Email: sayeadiiuc@gmail.com",
"article": 107
},
{
"id": 421,
"affiliation": [
{
"affiliation": "Department of Pharmacy, International Islamic University Chittagong, Chittagong-4318, Bangladesh"
}
],
"first_name": "Naureen",
"family_name": "Banu",
"email": null,
"author_order": 7,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 107
},
{
"id": 422,
"affiliation": [
{
"affiliation": "Department of Pharmacy, International Islamic University Chittagong, Chittagong-4318, Bangladesh"
}
],
"first_name": "Najmul",
"family_name": "Alam",
"email": null,
"author_order": 8,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 107
},
{
"id": 423,
"affiliation": [
{
"affiliation": "Department of Pharmacy, International Islamic University Chittagong, Chittagong-4318, Bangladesh"
}
],
"first_name": "Shahenur Alam",
"family_name": "Sakib",
"email": "sakibhasaniiuc@gmail.com",
"author_order": 9,
"ORCID": null,
"corresponding": true,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "Shahenur Alam Sakib, Department of Pharmacy, International Islamic University Chittagong, Chittagong-4318, Bangladesh, E-mail: sakibhasaniiuc@gmail.com",
"article": 107
}
],
"views": 1181,
"downloads": 110,
"references": [
{
"id": 3353,
"serial_number": 1,
"pmc": null,
"reference": "Costello EJ, Egger H, Angold A. 10-year research update review: the epidemiology of child and adolescent psychiatric disorders: I. Methods and public health burden. Journal of the American Academy of Child & Adolescent Psychiatry. 2005;44:972-86.",
"DOI": null,
"article": 107
},
{
"id": 3354,
"serial_number": 2,
"pmc": null,
"reference": "Costello EJ, Foley DL, Angold A. 10-year research update review: the epidemiology of child and adolescent psychiatric disorders: II. Developmental epidemiology. Journal of the American Academy of Child & Adolescent Psychiatry. 2006;45:8-25.",
"DOI": null,
"article": 107
},
{
"id": 3355,
"serial_number": 3,
"pmc": null,
"reference": "Okasha A. Mental health in the Middle East: an Egyptian perspective. Clinical psychology review. 1999;19:917-33.",
"DOI": null,
"article": 107
},
{
"id": 3356,
"serial_number": 4,
"pmc": null,
"reference": "Baumann P. Pharmacokinetic-pharmacodynamic relationship of the selective serotonin reuptake inhibitors. Clinical Pharmacokinetics. 1996;31:444-69.",
"DOI": null,
"article": 107
},
{
"id": 3357,
"serial_number": 5,
"pmc": null,
"reference": "Lambert C, Berlin I, Lee T-L, Hee SW, Tan AS, Picard D, et al. A standardized transcutaneous electric acupoint stimulation for relieving tobacco urges in dependent smokers. Evidence-Based Complementary and Alternative Medicine. 2011;2011.",
"DOI": null,
"article": 107
},
{
"id": 3358,
"serial_number": 6,
"pmc": null,
"reference": "Celada P, Bortolozzi A, Artigas F. Serotonin 5-HT 1A receptors as targets for agents to treat psychiatric disorders: rationale and current status of research. CNS Drugs. 2013;27:703-16.",
"DOI": null,
"article": 107
},
{
"id": 3359,
"serial_number": 7,
"pmc": null,
"reference": "Kanba S. Although antidepressants and anxiolytics are frequently used together to treat depression in the acute phase, how effective is the concomitant use of these drugs? Journal of Psychiatry and Neuroscience. 2004;29:485-6.",
"DOI": null,
"article": 107
},
{
"id": 3360,
"serial_number": 8,
"pmc": null,
"reference": "Greenbaum DS, Mayle JE, Vanegeren LE, Jerome JA, Mayor JW, Greenbaum RB, et al. Effects of desipramine on irritable bowel syndrome compared with atropine and placebo. Digestive diseases and sciences. 1987;32:257-66.",
"DOI": null,
"article": 107
},
{
"id": 3361,
"serial_number": 9,
"pmc": null,
"reference": "Novokhatny V, Taylor K, Zimmerman T. Thrombolytic potency of acid‐stabilized plasmin: superiority over tissue‐type plasminogen activator in an in vitro model of catheter‐assisted thrombolysis. Journal of Thrombosis and Haemostasis. 2003;1:1034-41.",
"DOI": null,
"article": 107
},
{
"id": 3362,
"serial_number": 10,
"pmc": null,
"reference": "Banerjee A, Chisti Y, Banerjee U. Streptokinase—a clinically useful thrombolytic agent. Biotechnology advances. 2004;22:287-307.",
"DOI": null,
"article": 107
},
{
"id": 3363,
"serial_number": 11,
"pmc": null,
"reference": "Thapar N, Sanderson IR. Diarrhoea in children: an interface between developing and developed countries. The lancet. 2004;363:641-53.",
"DOI": null,
"article": 107
},
{
"id": 3364,
"serial_number": 12,
"pmc": null,
"reference": "Silveira D, de Melo AF, Magalhães P, Fonseca-Bazzo Y. Tabernaemontana species: Promising sources of new useful drugs. Studies in Natural Products Chemistry: Elsevier; 2017. p. 227-89.",
"DOI": null,
"article": 107
},
{
"id": 3365,
"serial_number": 13,
"pmc": null,
"reference": "Slinkard K, Singleton VL. Total phenol analysis: automation and comparison with manual methods. American journal of enology and viticulture. 1977;28:49-55.",
"DOI": null,
"article": 107
},
{
"id": 3366,
"serial_number": 14,
"pmc": null,
"reference": "Charrouf Z, Guillaume D. Ethnoeconomical, ethnomedical, and phytochemical study of Argania spinosa (L.) Skeels. Journal of Ethnopharmacology. 1999;67:7-14.",
"DOI": null,
"article": 107
},
{
"id": 3367,
"serial_number": 15,
"pmc": null,
"reference": "Costa APR, Vieira C, Bohner LO, Silva CF, da Silva Santos EC, De Lima TCM, et al. A proposal for refining the forced swim test in Swiss mice. Progress in Neuro-Psychopharmacology and Biological Psychiatry. 2013;45:150-5.",
"DOI": null,
"article": 107
},
{
"id": 3368,
"serial_number": 16,
"pmc": null,
"reference": "de Almeida RN, Motta SC, de Brito Faturi C, Catallani B, Leite JR. Anxiolytic-like effects of rose oil inhalation on the elevated plus-maze test in rats. Pharmacology Biochemistry and Behavior. 2004;77:361-4.",
"DOI": null,
"article": 107
},
{
"id": 3369,
"serial_number": 17,
"pmc": null,
"reference": "Rabbani M, Sajjadi S, Zarei H. Anxiolytic effects of Stachys lavandulifolia Vahl on the elevated plus-maze model of anxiety in mice. Journal of Ethnopharmacology. 2003;89:271-6.",
"DOI": null,
"article": 107
},
{
"id": 3370,
"serial_number": 18,
"pmc": null,
"reference": "Sonavane G, Sarveiya V, Kasture V, Kasture S. Anxiogenic activity of Myristica fragrans seeds. Pharmacology Biochemistry and Behavior. 2002;71:239-44.",
"DOI": null,
"article": 107
},
{
"id": 3371,
"serial_number": 19,
"pmc": null,
"reference": "Chopin P, Pellow S, File SE. The effects of yohimbine on exploratory and locomotor behaviour are attributable to its effects at noradrenaline and not at benzodiazepine receptors. Neuropharmacology. 1986;25:53-7.",
"DOI": null,
"article": 107
},
{
"id": 3372,
"serial_number": 20,
"pmc": null,
"reference": "Borsini F, Meli A. Is the forced swimming test a suitable model for revealing antidepressant activity? Psychopharmacology. 1988;94:147-60.",
"DOI": null,
"article": 107
},
{
"id": 3373,
"serial_number": 21,
"pmc": null,
"reference": "Detke MJ, Rickels M, Lucki I. Active behaviors in the rat forced swimming test differentially produced by serotonergic and noradrenergic antidepressants. Psychopharmacology. 1995;121:66-72.",
"DOI": null,
"article": 107
},
{
"id": 3374,
"serial_number": 22,
"pmc": null,
"reference": "Cryan JF, Mombereau C, Vassout A. The tail suspension test as a model for assessing antidepressant activity: review of pharmacological and genetic studies in mice. Neuroscience & Biobehavioral Reviews. 2005;29:571-625.",
"DOI": null,
"article": 107
},
{
"id": 3375,
"serial_number": 23,
"pmc": null,
"reference": "Karolewicz B, Paul IA. Group housing of mice increases immobility and antidepressant sensitivity in the forced swim and tail suspension tests. European Journal of Pharmacology. 2001;415:197-201.",
"DOI": null,
"article": 107
},
{
"id": 3376,
"serial_number": 24,
"pmc": null,
"reference": "Sikder MAA, Siddique AB, Hossian A, Miah MK, Kaisar MA, Rashid MA. Evaluation of thrombolytic activity of four Bangladeshi medicinal plants, as a possible renewable source for thrombolytic compounds. Journal of Pharmacy and Nutrition Sciences. 2011;1:4-8.",
"DOI": null,
"article": 107
},
{
"id": 3377,
"serial_number": 25,
"pmc": null,
"reference": "Uddin MMN, Shahriar M. Phytochemical screenings, thrombolytic activity and antimicrobial properties of the bark extracts of Averrhoa bilimbi. Journal of Applied Pharmaceutical Science. 2013;3:094-6.",
"DOI": null,
"article": 107
},
{
"id": 3378,
"serial_number": 26,
"pmc": null,
"reference": "Atanasov AG, Waltenberger B, Pferschy-Wenzig E-M, Linder T, Wawrosch C, Uhrin P, et al. Discovery and resupply of pharmacologically active plant-derived natural products: A review. Biotechnology advances. 2015;33:1582-614.",
"DOI": null,
"article": 107
},
{
"id": 3379,
"serial_number": 27,
"pmc": null,
"reference": "Majid M, Khan MR, Shah NA, Haq IU, Farooq MA, Ullah S, et al. Studies on phytochemical, antioxidant, anti-inflammatory and analgesic activities of Euphorbia dracunculoides. BMC complementary and alternative medicine. 2015;15:349.",
"DOI": null,
"article": 107
},
{
"id": 3380,
"serial_number": 28,
"pmc": null,
"reference": "Wadood A, Ghufran M, Jamal SB, Naeem M, Khan A, Ghaffar R. Asnad. Phytochemical analysis of medicinal plants occurring in local area of Mardan Biochem Anal Biochem. 2013;2:1-4.",
"DOI": null,
"article": 107
},
{
"id": 3381,
"serial_number": 29,
"pmc": null,
"reference": "Zellweger R, Ayala A, DeMaso CM, Chaudry IH. Trauma-hemorrhage causes prolonged depression in cellular immunity. Shock. 1995;4:149-53.",
"DOI": null,
"article": 107
},
{
"id": 3382,
"serial_number": 30,
"pmc": null,
"reference": "Lauber C, Falcato L, Nordt C, Rössler W. Lay beliefs about causes of depression. Acta Psychiatrica Scandinavica. 2003;108:96-9.",
"DOI": null,
"article": 107
},
{
"id": 3383,
"serial_number": 31,
"pmc": null,
"reference": "Peng W-H, Lo K-L, Lee Y-H, Hung T-H, Lin Y-C. Berberine produces antidepressant-like effects in the forced swim test and in the tail suspension test in mice. Life Sciences. 2007;81:933-8.",
"DOI": null,
"article": 107
},
{
"id": 3384,
"serial_number": 32,
"pmc": null,
"reference": "Steru L, Chermat R, Thierry B, Simon P. The tail suspension test: a new method for screening antidepressants in mice. Psychopharmacology. 1985;85:367-70.",
"DOI": null,
"article": 107
},
{
"id": 3385,
"serial_number": 33,
"pmc": null,
"reference": "Page ME, Detke MJ, Dalvi A, Kirby LG, Lucki I. Serotonergic mediation of the effects of fluoxetine, but not desipramine, in the rat forced swimming test. Psychopharmacology. 1999;147:162-7.",
"DOI": null,
"article": 107
},
{
"id": 3386,
"serial_number": 34,
"pmc": null,
"reference": "Borsini F, Volterra G, Meli A. Does the behavioral “despair” test measure “despair”? Physiology & behavior. 1986;38:385-6.",
"DOI": null,
"article": 107
},
{
"id": 3387,
"serial_number": 35,
"pmc": null,
"reference": "Porsolt R, Bertin A, Jalfre M. Behavioral despair in mice: a primary screening test for antidepressants. Archives internationales de pharmacodynamie et de therapie. 1977;229:327.",
"DOI": null,
"article": 107
},
{
"id": 3409,
"serial_number": 36,
"pmc": null,
"reference": "Huffman JC, Smith FA, Blais MA, Januzzi JL, Fricchione GL. Anxiety, independent of depressive symptoms, is associated with in-hospital cardiac complications after acute myocardial infarction. Journal of psychosomatic research. 2008;65:557-63.",
"DOI": null,
"article": 107
},
{
"id": 3410,
"serial_number": 37,
"pmc": null,
"reference": "Hasanat A, Kabir MSH, Ansari MA, Chowdhury TA, Hossain MM, Islam MN, et al. Ficus cunia Buch.-Ham. ex Roxb.(leaves): An experimental evaluation of the cytotoxicity, thrombolytic, analgesic and neuropharmacological activities of its methanol extract. Journal of Basic and Clinical Physiology and Pharmacology. 2019;30.",
"DOI": null,
"article": 107
},
{
"id": 3411,
"serial_number": 38,
"pmc": null,
"reference": "Brown E, Hurd NS, McCall S, Ceremuga TE. Evaluation of the anxiolytic effects of chrysin, a Passiflora incarnata extract, in the laboratory rat. AANA journal. 2007;75.",
"DOI": null,
"article": 107
},
{
"id": 3412,
"serial_number": 39,
"pmc": null,
"reference": "Rickels K, Case WG, Downing RW, Winokur A. Long-term diazepam therapy and clinical outcome. Jama. 1983;250:767-71.",
"DOI": null,
"article": 107
},
{
"id": 3413,
"serial_number": 40,
"pmc": null,
"reference": "Bodnoff SR, Suranyi-Cadotte B, Aitken DH, Quirion R, Meaney MJ. The effects of chronic antidepressant treatment in an animal model of anxiety. Psychopharmacology. 1988;95:298-302.",
"DOI": null,
"article": 107
},
{
"id": 3414,
"serial_number": 41,
"pmc": null,
"reference": "Brown GR, Nemes C. The exploratory behaviour of rats in the hole-board apparatus: is head-dipping a valid measure of neophilia? Behavioural processes. 2008;78:442-8.",
"DOI": null,
"article": 107
},
{
"id": 3415,
"serial_number": 42,
"pmc": null,
"reference": "Crawley J, Davis L. Baseline exploratory activity predicts anxiolytic responsiveness to diazepam in five mouse strains. Brain research bulletin. 1982;8:609-12.",
"DOI": null,
"article": 107
},
{
"id": 3416,
"serial_number": 43,
"pmc": null,
"reference": "Calabrese EJ. An assessment of anxiolytic drug screening tests: hormetic dose responses predominate. Critical reviews in toxicology. 2008;38:489-542.",
"DOI": null,
"article": 107
},
{
"id": 3417,
"serial_number": 44,
"pmc": null,
"reference": "Nolan NA, Parkes M. The effects of benzodiazepines on the behaviour of mice on a hole-board. Psychopharmacologia. 1973;29:277-88.",
"DOI": null,
"article": 107
},
{
"id": 3418,
"serial_number": 45,
"pmc": null,
"reference": "Lourenço da Silva A, Elisabetsky E. Interference of propylene glycol with the hole-board test. Brazilian Journal of Medical and Biological Research. 2001;34:545-7.",
"DOI": null,
"article": 107
},
{
"id": 3419,
"serial_number": 46,
"pmc": null,
"reference": "Kaur R, Kaur S. Anxiolytic potential of methanol extract from Ageratum conyzoides Linn Leaves. Pharmacognosy Journal. 2015;7.",
"DOI": null,
"article": 107
},
{
"id": 3420,
"serial_number": 47,
"pmc": null,
"reference": "Brogden R, Speight T, Avery G. Streptokinase: a review of its clinical pharmacology, mechanism of action and therapeutic uses. Drugs. 1973;5:357-445.",
"DOI": null,
"article": 107
},
{
"id": 3421,
"serial_number": 48,
"pmc": null,
"reference": "Shifah F, Tareq AM, Sayeed MA, Islam MN, Emran TB, Ullah MA, et al. Antidiarrheal, cytotoxic and thrombolytic activities of methanolic extract of Hedychium coccineum leaves. measurement.11:12.",
"DOI": null,
"article": 107
},
{
"id": 3422,
"serial_number": 49,
"pmc": null,
"reference": "Taylor FB, MÜLLER-EBERHARD HJ. Factors influencing lysis of whole blood clots. Nature. 1967;216:1023-5.",
"DOI": null,
"article": 107
},
{
"id": 3423,
"serial_number": 50,
"pmc": null,
"reference": "Korninger C, Collen D. Studies on the specific fibrinolytic effect of human extrinsic (tissue-type) plasminogen activator in human blood and in various animal species in vitro. Thrombosis and haemostasis. 1981;46:561-5.",
"DOI": null,
"article": 107
},
{
"id": 3424,
"serial_number": 51,
"pmc": null,
"reference": "Ambrus CM, Back N, Ambrus JL. On the mechanism of thrombolysis by plasmin. Circulation research. 1962;10:161-5.",
"DOI": null,
"article": 107
},
{
"id": 3425,
"serial_number": 52,
"pmc": null,
"reference": "Gryglewski RJ, Korbut R, Robak J, Świȩs J. On the mechanism of antithrombotic action of flavonoids. Biochemical pharmacology. 1987;36:317-22.",
"DOI": null,
"article": 107
},
{
"id": 3426,
"serial_number": 53,
"pmc": null,
"reference": "Bhowmick R, Sarwar MS, RahmanDewan SM, Das A, Das B, NasirUddin MM, et al. In vivo analgesic, antipyretic, and anti-inflammatory potential in Swiss albino mice and in vitro thrombolytic activity of hydroalcoholic extract from Litsea glutinosa leaves. Biological Research. 2014;47:56.",
"DOI": null,
"article": 107
}
]
},
{
"id": 105,
"slug": "178-1595827549-adaptations-of-muscular-biology-in-response-to-potential-glucocorticoid-treatment-in-broiler-chicken",
"featured": false,
"slider": false,
"issue": "Vol4 Issue1",
"type": "original_article",
"manuscript_id": "178-1595827549",
"recieved": "2020-06-17",
"revised": null,
"accepted": "2020-08-31",
"published": "2020-09-13",
"pdf_file": "https://jabet.bsmiab.org/media/pdf_file/2023/51/178-1595827549.pdf",
"title": "Adaptations of muscular biology in response to potential glucocorticoid treatment in broiler chicken",
"abstract": "<p>Poultry meat production has been dramatically increased in the last few decades due to increased population rate. Glucocorticoids decrease the growth of poultry and increase fat accumulation in liver and meat. In the coming days, it is important to consider the quality of meat to fulfill the increasing demand of proteins. The morphological and biometric properties of meat are associated with the quality of meat. The present research aimed to study the adaptations of muscular biology in response to potential glucocorticoid treatment in broiler chicken. This experiment was conducted into three groups of broilers (i.e. control group: homemade ration, group A: commercial broiler ration, and group B: a high dose of glucocorticoid at 7 mg/kg) started from day 7 to 28. Meat and blood samples were collected at day 7, 14, 21, and 28. For gross morphology, color and weight of meat were measured. Histomorphology of meat was studied under light microscope by Hematoxylin & Eosin stain. The length and width of meat fibers were measured using calibrated stage micrometer. The blood cholesterol dynamics was measured by spectrophotometer. The color of breast meat was more yellowish and lighter than thigh meat. The weight of meat was negatively affected by glucocorticoid. Glucocorticoid treatment negatively influenced the number of myofibers in breast meat, while positively influenced the thigh meat. Excess dietary glucocorticoid increased the biometry of breast meat and decreased that of thigh meat in broiler. Glucocorticoid non-significantly increased the serum cholesterol level. These findings advance our knowledge about the action of glucocorticoid in the muscular system and provide basis for novel therapies to prevent glucocorticoid-induced muscular atrophy.</p>",
"journal_reference": "J Adv Biotechnol Exp Ther. 2021; 4(1): 01-08.",
"academic_editor": "Md Nabiul Islam, PhD; Yamaguchi University, Japan",
"cite_info": "Akter F, Sultana N, et al. Adaptations of muscular biology in response to potential glucocorticoid treatment in broiler chicken. J Adv Biotechnol Exp Ther. 2021; 4(1): 01-08.",
"keywords": [
"Morphology",
"Muscular biology",
"Glucocorticoid",
"Broilers",
"Meat"
],
"DOI": "10.5455/jabet.2021.d100",
"sections": [
{
"section_number": 1,
"section_title": "INTRODUCTION",
"body": "<p>Due to the growing human population and increasing demand for proteins throughout the world, the production of poultry meat has dramatically increased in the last few decades. A broiler can attain a body weight of 2 kg by consuming 3 kg of feed within 35 days [<a href=\"#r-1\">1</a>]. The majority of this increase has been rendered possible due to rapid growth by genetic selection of poultry for quantitative traits, which has tremendously augmented the growth rate of meat by inducing hypertrophy in meat fibers [<a href=\"#r-2\">2, 3</a>]. In the coming decades, it is also essential to ensure the quality meat to fulfill the increasing demand of proteins. The synthetic steroid, glucocorticoid is used as a growth promoting agents for increasing the body weight in livestock legally or illegally [<a href=\"#r-4\">4, 5, 6</a>]. Steroid hormones are applied for increasing the growth rate of broiler to meet up the demand of total meat consumption [<a href=\"#r-7\">7</a>].<br />\r\nThe color of meat is an important quality attributes both for the consumer’s selection of fresh meat at the retail level and for the consumer’s final evaluation and acceptance of meat product [<a href=\"#r-8\">8</a>]. Morphological and biometric properties of meat fibers play a key role in performance and meat quality in broiler chicken [<a href=\"#r-9\">9</a>]. So, investigation of these characteristics is one of the most practical importance to poultry and meat scientists. Although the current commercial broiler chicken strains are a result of successful selection programs for rapid growth and body conformation, especially favoring the breast muscles. Because, the breast meat is the most valuable portion of the chicken meat in the market and have significant economic impact. For this reason, the broiler industry is constantly interested in evaluating the weight and yield of the breast meat as the most important variables [<a href=\"#r-10\">10</a>].<br />\r\nIt has been reported that dietary supplementation of steroid growth promoter; glucocorticoid did not improve overall growth performance (i.e. body weight and dressed weight) of broiler chicken [<a href=\"#r-11\">11</a>]. In chicken, glucocorticoid (dexamethasone) induced-retarded fatty acid utilization in muscle. Dietary dexamethasone promoted the transcriptional activity of genes related to lipid uptake and oxidation in muscles. Unrivalled lipid uptake and utilization are recommended to be involved in the augmented intramyocellular lipid accumulation [<a href=\"#r-12\">12</a>]. A higher dose of glucocorticoid causes greater muscular atrophy in rat in comparison to those receiving lower doses of glucocorticoid [<a href=\"#r-13\">13, 14</a>]. But the mechanism underlying the muscular atrophy in response to glucocorticoid in broiler chicken is not completely understood. It was hypothesized that high dose of glucocorticoid alters the number or the size of myofibers in broiler chicken.<br />\r\nIn addition, modern intensive broiler production has been in the efficient and economical production of high-quality chicken meat [<a href=\"#r-15\">15</a>]. The quality of the meat is associated with the total lipid and cholesterol profile in blood. A group of scientists have noted that supplementation of herbal mixture lowers the blood cholesterol level in broiler chicken [<a href=\"#r-15\">15</a>]. There is no research exists which studies the effects of dexamethasone on the blood cholesterol profile in broiler chickens. Therefore, the present study aimed to study the morphology and biometry of breast and thigh meat and on blood cholesterol level in response to high dose of glucocorticoid in broiler chicken.</p>"
},
{
"section_number": 2,
"section_title": "MATERIALS AND METHODS",
"body": "<p><strong>Animal ethics</strong><br />\r\nThe present experimental procedures were approved and performed in accordance with the guidelines for the care and use of animals as established by Animal Welfare and Experimentation Ethics Committee, Bangladesh Agricultural University, Bangladesh.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Bird management</strong><br />\r\nThe experiment was conducted in the department of Anatomy and Histology, Bangladesh Agricultural University, Mymensingh-2202. In total, 90 (“Cobb 500” strain) one day old broiler chicks were purchased from a commercial hatchery (Provita Feed and Hatchery Ltd., Bangladesh). Upon arrival, broilers chicks were individually weighed, and steel wing tagged before being allotted to one of the 3 pens (thirty birds per pen), equally distributed over three identical climate-controlled rooms, so that each pen had a similar initial total body weight. Each pen had three drinking nipples with a cup underneath connected to a water tank. A feeding tray was placed on the floor of the pen during week 1 and replaced by a feeding trough thereafter. Feed and water were available ad libitum throughout the experiment. Rice husk was used as litter material.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Experimental design and treatments</strong><br />\r\nThe experiment was conducted into three groups in three different pens (three pens with thirty birds per pen), and treatments were equally distributed over pens. All birds received the same standard starter diet for the first 7 days of the experiment. After week 1, three dietary treatments were provided which contained homemade ration in control group, commercial broiler ration in group A and homemade ration with a high dose of glucocorticoid (dexamethasone) (7mg/kg) in group B. The homemade ration was formulated according to the commercial broiler type ration to meet the nutrient recommendations for boilers. These dietary treatments were randomly assigned to the broilers in pens. All rations were provided as pellets for the starter and grower diet. The broiler ration did not contain any antibiotic or other growth promoters.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Tissue collection and preservation</strong><br />\r\nThe experimental broilers were slaughtered by cervical subluxation method at day 7, 14, 21, and 28 of the experiment. The birds were plucked immediately after slaughtering and meat samples were excised from the breast and thigh of the broilers, rinsed with physiological saline (0.9%). Then the collected meat samples were immediately placed in Bouin’s solution for further histological study.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Blood collection and serum preparation</strong><br />\r\nBlood samples were collected from wing vein of each broiler at day 7, 14, 21 and 28 days of the experiment. About 5 ml of blood was collected from each broiler without anticoagulant, in a sterile glass test tube. The blood containing tubes were placed in a slanting position at room temperature for clotting. The tubes were then placed in a refrigerator at 4ºC over-night. Serum sample was then collected and centrifuged at 1000 rpm for 15 minutes to get rid of unwanted substances. The collected serum was then stored in a screw capped serum vial and preserved at -20º C until use.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Gross morphology</strong><br />\r\nMorphometric (i.e. color and weight) alterations of the breast and thigh meat were examined in the broilers during the experiment. The color of the excised meat samples was compared in between breast and thigh meat through visual inspection. The weight of breast and thigh meat was recorded by electronic digital weighing balance.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Histomorphology and biometric measurements</strong><br />\r\nFor histological studies, the collected breast and thigh meat were dehydrated in the series of ascending graded alcohol followed by clearing in three changes of xylene. Then, the tissues were infiltrated with different grades of melted paraffin (49°C, 55°C, and 58°C) at 30 minutes interval. The tissues were then embedded in paraffin (58°C) and finally the sections were cut at 6μ thickness using sliding microtome (MIC 509, Euromex, Japan). After cutting, the sections were floated in a floatation bath at 37°C for stretching. Then the sections were mounted on clean slides using an adhesive (egg albumins) and dried on a slide warmer at 37°C. The sections were then stained using Mayer’s Hematoxylin and Eosin (H & E) stain. The histological characters (i.e. number of meat fibers) of meat tissues were observed under light microscope at 40x magnification. The length and width of breast and thigh meat fibers were considered for biometric measurement which was performed using calibrated stage micrometer in μm (micrometer). Thirty sections (ten sections from each group) were biometrically evaluated from three different groups of experiment.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Photomicrographs</strong><br />\r\nThe sections were evaluated at 400-fold magnification by light microscope (Leica DMR; Leica Microsystems, Wetzlar, Germany). The pictures were taken with a digital color camera. For each section, 10 randomly selected fields were captured for the measurement of number, length and width of fibers, as well as the total number of fibers. Results are presented as the mean percentage of the fibers per total fibers in the evaluated fields.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Serum cholesterol analysis</strong><br />\r\n5 ml of blood was collected without anticoagulant in sterile glass test tubes. The blood containing tubes were placed in a slanting position at room temperature for clotting. The tubes were then placed in the refrigerator at 4<sup>0</sup>C over-night. Blood was centrifuged at 1000 rpm for 15 minutes for serum collection and the serum was stored in a freezer at -20°C until analyses. Blood serum cholesterol (TCHOL) was determined by the spectrophotometric method, using Human Humalyzer 2000 (Wiesbaden, Germany). The cholesterol present in the serum sample produced a colored complex. The intensity of the color formed is proportional to the cholesterol concentration in the serum sample.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Statistical analysis</strong><br />\r\nThe results were analyzed in the GraphPad Prism version 8.0 (GraphPad Software, San Diego, CA, USA). All data were expressed as mean ± standard error and differences among the groups of birds were compared using one-way ANOVA with post-hoc Duncan’s multiple range test. Values of p<0.05 were considered significant.</p>"
},
{
"section_number": 3,
"section_title": "RESULTS",
"body": "<p><strong>Gross morphology of breast and thigh meat</strong><br />\r\nThe color and lightness of breast and thigh meat were observed by visual inspection. The breast meat exhibited a light-yellow color, while thigh meat appeared pale red in the broilers of group B (<a href=\"#figure1\">Figure 1a</a>). The color of breast meat was more yellowish as compared to thigh meat. The lightness of breast meat was more than the thigh meat.<br />\r\nThe effects of glucocorticoid on the weight gain of breast and thigh meat in broiler chicken are presented in<a href=\"#figure2\"> Figure 2</a>. The breast meat weight was decreased in group B as compared to control and group A in broiler at day 21 of the experiment. The thigh meat weight was decreased in the broilers of group B than the broilers of control and group A at days 21 and 28 of the experiment.</p>\r\n\r\n<div id=\"figure1\">\r\n<figure class=\"image\"><img alt=\"\" height=\"412\" src=\"/media/article_images/2024/38/03/178-1595827549-Figure1.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 1.</strong> a. Representative photos of breast and thigh meat color of broilers in group B at day 14 of the experiment. b. Histological representation of breast and thigh meat fibers (40X). Bm- breast meat, tm- thigh meat.</figcaption>\r\n</figure>\r\n</div>\r\n\r\n<div id=\"figure2\">\r\n<figure class=\"image\"><img alt=\"\" height=\"567\" src=\"/media/article_images/2024/38/03/178-1595827549-Figure2.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 2. </strong>Effects of glucocorticoid on the weight of breast and thigh meat (g) in relation to the days of the experiment in broiler. Data were expressed as mean ± standard error and differences among the groups of birds were compared using one-way ANOVA with post-hoc Duncan’s multiple range test. Column with differing superscripts are significantly different from each other (P < 0.05).</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Histology of breast and thigh meat</strong><br />\r\nThe histological structures of breast and thigh meat fibers are presented in broiler chicken (<a href=\"#figure1\">Figure 1b</a>). Steroid growth promoter, glucocorticoid influenced the numbers of myofibers in breast and thigh meat of broiler chicken (<a href=\"#figure3\">Figure 3</a>). The number of myofibers of breast meat was decreased in the broilers of group A and B as compared to the control birds at day 21 of the experiment. The number of myofibers of thigh meat was increased in the broiler of group B as compared to that of group A and control at day 7, 14, 21, and 28 of the experiment.</p>\r\n\r\n<div id=\"figure3\">\r\n<figure class=\"image\"><img alt=\"\" height=\"567\" src=\"/media/article_images/2024/38/03/178-1595827549-Figure3.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 3. </strong>Effects of glucocorticoid on the number of breast and thigh meat fibers in relation to days of the experiment in broiler chicken. Data were expressed as mean ± standard error and differences among the groups of birds were compared using one-way ANOVA with post-hoc Duncan’s multiple range test. Column with differing superscripts are significantly different from each other (P < 0.05).</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Biometry of breast and thigh meat</strong><br />\r\nThe biometry (i.e. length and width of myofibers) of breast and thigh meat fibers is affected by dietary glucocorticoid in broiler (<a href=\"#figure4\">Figure 4</a> & <a href=\"#figure5\">5</a>). The length of breast meat fiber was non significantly increased in group B as compared to the control birds, while the length of thigh meat fibers was significantly decreased in the broilers of group B as compared to control and group A at day 7, 21, and 28 of the experiment. The width of breast meat fiber was non-significantly increased in group B as compared to the control birds, while the width of thigh meat fibers was significantly decreased in the broiler of group B as compared to control and group A at day 14 and 28 of the experiment.</p>\r\n\r\n<div id=\"figure4\">\r\n<figure class=\"image\"><img alt=\"\" height=\"621\" src=\"/media/article_images/2024/38/03/178-1595827549-Figure4.jpg\" width=\"469\" />\r\n<figcaption><strong>Figure 4. </strong>Effects of glucocorticoid on the length of breast and thigh meat fibers (µm) in relation to the days of the experiment in broiler chicken. Data were expressed as mean ± standard error and differences among the groups of birds were compared using one-way ANOVA with post-hoc Duncan’s multiple range test. Column with differing superscripts are significantly different from each other (P < 0.05).</figcaption>\r\n</figure>\r\n</div>\r\n\r\n<div id=\"figure5\">\r\n<figure class=\"image\"><img alt=\"\" height=\"597\" src=\"/media/article_images/2024/38/03/178-1595827549-Figure5.jpg\" width=\"477\" />\r\n<figcaption><strong>Figure 5.</strong> Effects of glucocorticoid in the width of breast and thigh meat fibers (µm) in relation to the days of the experiment in broiler chicken. Data were expressed as mean ± standard error and differences among the groups of birds were compared using one-way ANOVA with post-hoc Duncan’s multiple range test. Column with differing superscripts are significantly different from each other (P < 0.05).</figcaption>\r\n</figure>\r\n</div>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Blood cholesterol profile</strong><br />\r\nThe serum cholesterol data in different groups of broilers are presented in <a href=\"#figure6\">Figure 6</a>. There was a non-significant increase in serum cholesterol level in broilers of group B as compared to control and group A at day 7, 21, and 28 of the experiment.</p>\r\n\r\n<div id=\"figure6\">\r\n<figure class=\"image\"><img alt=\"\" height=\"341\" src=\"/media/article_images/2024/38/03/178-1595827549-Figure6.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 6. </strong>Dynamics of cholesterol in serum in relation to age of broiler chicken treated with glucocorticoid. Data were expressed as mean ± standard error and differences among the groups of birds were compared using one-way ANOVA with post-hoc Duncan’s multiple range test.</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>"
},
{
"section_number": 4,
"section_title": "DISCUSSION",
"body": "<p>The present study intended to demonstrate the effects of glucocorticoid on gross morphology (i.e. color and weight), histomorphology (i.e. number of myofibers), and biometry (i.e. length and width of myofibers) of breast and thigh meat and blood cholesterol level in broiler chicken. It was hypothesized that high dose of glucocorticoid would result alteration in the morphology of meat, which coincides with reduced body weight. Additionally, it was questioned that the high dose of glucocorticoid could alleviate the blood level of cholesterol in broiler. Therefore, the experiment was conducted with high dose of glucocorticoid in broiler chicken.<br />\r\nThe color of meat is an important quality attribute for the consumer’s selection of fresh meat and acceptance of a meat product [<a href=\"#r-8\">8</a>]. Extreme paleness or darkness of meat has reflected badly on poultry industry [<a href=\"#r-16\">16</a>]. The color of poultry meat is highly correlated to the amount of haem containing compounds, myoglobin. However, within most individual experiments the myoglobin content in breast muscle was significantly lower than that of the thigh muscle. The thigh meat has a high proportion of myoglobin as compared to breast meat [<a href=\"#r-17\">17</a>]. The intramuscular connective tissue and tenderness of the meat are varied between the breast and thigh meat in chicken [<a href=\"#r-18\">18</a>]. The color of breast meat is more yellowish as compared to the thigh meat. The lightness of breast meat is more than the thigh. The variations in color and lightness might be related with myoglobin content in meat, the intramuscular connective tissues and tenderness of the meat.<br />\r\nHigher doses of dexamethasone have negative effects on the growth rate and tibia genomic properties of broiler [<a href=\"#r-19\">19</a>]. The immunosuppressive effect of glucocorticoid is related with the muscle fiber loss in muscular dystrophy [<a href=\"#r-20\">20</a>]. In addition, glucocorticoid impairs young myofibers formation in skeletal muscle, increase protein breakdown in skeletal muscle of young adults, decrease protein synthesis in aged rats, and morphologically and functionally damage muscle precursor cells [<a href=\"#r-21\">21</a>].<br />\r\nGlucocorticoid; dexamethasone increases the plasma T3 levels and metabolism of protein in muscle which is responsible for muscular dystrophy [<a href=\"#r-22\">22</a>]. The present study reported that dietary supplementation of glucocorticoid decreased the individual weight of breast and thigh meat in broiler chicken. These results support the study performed by our group and suggested that dietary glucocorticoid did not improve overall growth performance (i.e. body weight and dressed weight) in broiler chicken [<a href=\"#r-11\">11</a>]. This is in agreement with other researches [<a href=\"#r-14\">14</a>, <a href=\"#r-22\">22</a>] who reported that high doses of dexamethasone cause muscular dystrophy and atrophy in rat.<br />\r\nHistological and biochemical properties of meat fibers are associated with the meat quality. So, investigation of these characteristics is one of the most practical importance to poultry and meat scientists. However, still there is not enough research that definitely demonstrates the deleterious effects of increasing fiber size on meat quality, and some researchers showed that largest breast meat fibers exhibited high meat quality [<a href=\"#r-9\">9</a>]. In the present study, the number of myofibers of breast meat was decreased in the broiler fed the ration supplemented with dexamethasone as compared to the control. And the number of thigh meat fibers were increased in glucocorticoid treated broiler. So, it is assumed that the lower number of breast meat fibers might contribute to the reduced body weight gain in response to dietary glucocorticoid in broiler chicken.<br />\r\nMany researchers reported that excess glucocorticoid either endogenous or exogenous induce muscular atrophy leading to myopathy in skeletal muscle in animals [<a href=\"#r-13\">13</a>, <a href=\"#r-14\">14</a>]. Thus, high dose of glucocorticoid affects the quality of meat in animals. In addition, A high dose of glucocorticoid causes mild damage in the liver of rat and improves renal failure in multiple myeloma patients [<a href=\"#r-23\">23</a>, <a href=\"#r-24\">24</a>]. However, there is no study exists about the effects of high dose of glucocorticoid on the biometry of breast and thigh meat in broiler chicken. In this experiment, high dose of glucocorticoid affected the biometry of breast and thigh meat fiber in broiler chicken. The length of breast meat fibers was non significantly increased, while the length of thigh meat fibers was significantly decreased in the broilers. The width of breast meat fiber was non-significantly increased, while the width of thigh meat fibers was significantly decreased in the broiler. High dose of glucocorticoid has positive effects on the breast meat biometry and negative effects on the thigh meat biometric properties in broiler chicken. This result supports that the high dose of glucocorticoid reduces the myotube diameter in thigh meat [<a href=\"#r-25\">25</a>] but opposite to the breast meat profile in broiler.<br />\r\nThe cholesterol content in broiler meat is one of the quality indices whose level the consumer wants to have as less as possible. The ingredients of feed are one of the most contributing factors to reduce the cholesterol level in meat in poultry [<a href=\"#r-26\">26, 27</a>]. In the present study, dietary glucocorticoid was used in broiler and their influence on blood cholesterol dynamics was evaluated in the broiler chicken. In this study, the blood cholesterol level was non significantly increased in response to dietary glucocorticoid in broiler chicken. On the contrary, feed supplementation of herbal mixture lowers the blood cholesterol level in broiler chicken [<a href=\"#r-15\">15</a>]. This increased blood cholesterol might release into the body tissues i.e. meat and liver which is necessary to investigate as further research.<br />\r\nIn conclusion, the outcomes of the present study reveal that a high dose of glucocorticoid alters the breast and thigh meat morphology in broiler chickens. The reduced number of breast meat fibers might contribute to the decreased body weight gain in broiler treated with glucocorticoid. A high dose of glucocorticoid increased the breast meat biometry and decreased the thigh meat biometric properties in broiler chicken. The effects of glucocorticoid on the biometry of breast and thigh meat fibers might be associated with the excess of glucocorticoid in breast and thigh meat. Morphological and biometric properties of breast and thigh meat fibers can provide beneficial information regarding the nature of mechanically deboned meat and its use in food products. The up regulated blood cholesterol by glucocorticoid might release into body tissues i.e. meat and liver which is necessary to investigate as further research. It would be interesting to assess the effects of glucocorticoid on intramuscular connective tissue, lipid profile and cholesterol level of meat in broiler.</p>"
},
{
"section_number": 5,
"section_title": "ACKNOWLEDGEMENT",
"body": "<p>Technicians of the department of Anatomy and Histology, Bangladesh Agricultural University, Bangladesh are thanked for their assistance in performing the experiment. The author, Dr. Nasrin Sultana is gratefully acknowledged to the funding of the University Grants Commission (UGC) of Bangladesh (Project No.: Life 11/2017).</p>"
},
{
"section_number": 6,
"section_title": "CONFLICTS OF INTEREST",
"body": "<p>There is no conflict of interest among the authors.</p>"
},
{
"section_number": 7,
"section_title": "AUTHOR CONTRIBUTIONS",
"body": "<p>FA and MA were involved in conception and design of the experiments. FA and RI contributed to perform the experiments. FA and AK analyzed data. NS contributed to drafting the article. MHS and NS contributed to revising it critically for important intellectual content.</p>"
}
],
"figures": [
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/38/03/178-1595827549-Figure1.jpg",
"caption": "Figure 1. a. Representative photos of breast and thigh meat color of broilers in group B at day 14 of the experiment. b. Histological representation of breast and thigh meat fibers (40X). Bm- breast meat, tm- thigh meat.",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/38/03/178-1595827549-Figure2.jpg",
"caption": "Figure 2. Effects of glucocorticoid on the weight of breast and thigh meat (g) in relation to the days of the experiment in broiler. Data were expressed as mean ± standard error and differences among the groups of birds were compared using one-way ANOVA with post-hoc Duncan’s multiple range test. Column with differing superscripts are significantly different from each other (P < 0.05).",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/38/03/178-1595827549-Figure3.jpg",
"caption": "Figure 3. Effects of glucocorticoid on the number of breast and thigh meat fibers in relation to days of the experiment in broiler chicken. Data were expressed as mean ± standard error and differences among the groups of birds were compared using one-way ANOVA with post-hoc Duncan’s multiple range test. Column with differing superscripts are significantly different from each other (P < 0.05).",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/38/03/178-1595827549-Figure4.jpg",
"caption": "Figure 4. Effects of glucocorticoid on the length of breast and thigh meat fibers (µm) in relation to the days of the experiment in broiler chicken. Data were expressed as mean ± standard error and differences among the groups of birds were compared using one-way ANOVA with post-hoc Duncan’s multiple range test. Column with differing superscripts are significantly different from each other (P < 0.05).",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/38/03/178-1595827549-Figure5.jpg",
"caption": "Figure 5. Effects of glucocorticoid in the width of breast and thigh meat fibers (µm) in relation to the days of the experiment in broiler chicken. Data were expressed as mean ± standard error and differences among the groups of birds were compared using one-way ANOVA with post-hoc Duncan’s multiple range test. Column with differing superscripts are significantly different from each other (P < 0.05).",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/38/03/178-1595827549-Figure6.jpg",
"caption": "Figure 6. Dynamics of cholesterol in serum in relation to age of broiler chicken treated with glucocorticoid. Data were expressed as mean ± standard error and differences among the groups of birds were compared using one-way ANOVA with post-hoc Duncan’s multiple range test.",
"featured": false
}
],
"authors": [
{
"id": 406,
"affiliation": [
{
"affiliation": "Department of Pharmacology, Bangladesh Agricultural University, Mymensingh-2202, Bangladesh"
}
],
"first_name": "Ferdoushi",
"family_name": "Akter",
"email": null,
"author_order": 1,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 105
},
{
"id": 407,
"affiliation": [
{
"affiliation": "Department of Anatomy and Histology, Bangladesh Agricultural University, Mymensingh-2202, Bangladesh"
}
],
"first_name": "Nasrin",
"family_name": "Sultana",
"email": "nasrin.sultana@bau.edu.bd",
"author_order": 2,
"ORCID": null,
"corresponding": true,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "Nasrin Sultana, PhD; Department. of Anatomy and Histology, Bangladesh\r\nAgricultural University, Mymensingh-2202, Bangladesh, E-mail: nasrin.sultana@bau.edu.bd",
"article": 105
},
{
"id": 408,
"affiliation": [
{
"affiliation": "Department of Life Science and Veterinary Medicine, Bangabandhu Sheikh Mujibur Rahman Science and Technology University, Gopalganj 8100, Bangladesh"
}
],
"first_name": "Marzia",
"family_name": "Afrose",
"email": null,
"author_order": 3,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 105
},
{
"id": 409,
"affiliation": [
{
"affiliation": "Department of Anatomy and Histology, Bangladesh Agricultural University, Mymensingh-2202, Bangladesh"
}
],
"first_name": "Alamgir",
"family_name": "Kabir",
"email": null,
"author_order": 4,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 105
},
{
"id": 410,
"affiliation": [
{
"affiliation": "Department of Anatomy and Histology, Bangladesh Agricultural University, Mymensingh-2202, Bangladesh"
}
],
"first_name": "Rafiqul",
"family_name": "Islam",
"email": null,
"author_order": 5,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 105
},
{
"id": 411,
"affiliation": [
{
"affiliation": "Department of Pharmacology, Bangladesh Agricultural University, Mymensingh-2202, Bangladesh"
}
],
"first_name": "Mahmudul Hasan",
"family_name": "Sikder",
"email": null,
"author_order": 6,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 105
}
],
"views": 1244,
"downloads": 181,
"references": [
{
"id": 3226,
"serial_number": 1,
"pmc": null,
"reference": "Choct M. Managing Gut Health through Nutrition. Bri Poult Sci 2009; 50: 9–15. 10.1080/00071660802538632.",
"DOI": null,
"article": 105
},
{
"id": 3227,
"serial_number": 2,
"pmc": null,
"reference": "Picard B, Lefaucheur L, Berri C and Duclos MJ. Muscle fibre ontogenesis in farm animal species. Reprod Nutri Dev 2002; 42:415–431.",
"DOI": null,
"article": 105
},
{
"id": 3228,
"serial_number": 3,
"pmc": null,
"reference": "Scheuermann GN, Bilgili SF, Tuzun S and Mulvaney DR. Comparison of chicken genotypes: myofiber number in pectoralis muscle and myostatin ontogeny. Poult Sci 2004; 83:1404–1412.",
"DOI": null,
"article": 105
},
{
"id": 3229,
"serial_number": 4,
"pmc": null,
"reference": "Jeong SH, Kang D, Lim MW, Kang CS, and Sung HJ. Risk assessment of growth hormones and antimicrobial residues in meat. Toxicol Res 2010; 26: 301.",
"DOI": null,
"article": 105
},
{
"id": 3230,
"serial_number": 5,
"pmc": null,
"reference": "Yuan Y, Xu C, Peng C, Jin Z, Chen W, and Liu L . Analytical methods for the detection of corticosteroids-residues in animal-derived foodstuffs. Crit Rev Anal Chem 2008; 38: 227-241.",
"DOI": null,
"article": 105
},
{
"id": 3231,
"serial_number": 6,
"pmc": null,
"reference": "Serratosa J, Blass A, Rigau B, Mongrell B, Rigau T, Tortades M, et al. Residues from veterinary medicinal products, growth promoters and performance enhancers in food-producing animals: a European Union perspective. Rev Sci Tech 2006; 25: 637-653.",
"DOI": null,
"article": 105
},
{
"id": 3232,
"serial_number": 7,
"pmc": null,
"reference": "Schumacher HI, Barrantes SQ, Alpízar MZ and Corella MR. Serum sexual steroid hormones and lipids in commercial broilers (Gallus domesticus) in Costa Rica. J Appl Poultry Res 2010; 19: 279–287.",
"DOI": null,
"article": 105
},
{
"id": 3233,
"serial_number": 8,
"pmc": null,
"reference": "Fletcher DL, QIAO M, and Smith DP. The Relationship of Raw Broiler Breast Meat Color and pH to Cooked Meat Color and pH. Poult Sci 2000; 79:784–788.",
"DOI": null,
"article": 105
},
{
"id": 3234,
"serial_number": 9,
"pmc": null,
"reference": "Tůmová E and Teimouri A. Chicken muscle fibres characteristics and meat quality: a review. Sci Agric bohem 2009; 40 : 253–258.",
"DOI": null,
"article": 105
},
{
"id": 3235,
"serial_number": 10,
"pmc": null,
"reference": "Scheuermann GN, Bilgili SF, Hess JB and Mulvaney DR. Breast muscle development in commercial broiler chickens. Poult Sci 2003; 82:1648-1658.",
"DOI": null,
"article": 105
},
{
"id": 3236,
"serial_number": 11,
"pmc": null,
"reference": "Afrose M, Sultana N, and Islam MR. Physiological Responses of Corticosteroid, Dexamethasone in Broiler Chicken. IJSR 2018; 7:1459-63.",
"DOI": null,
"article": 105
},
{
"id": 3237,
"serial_number": 12,
"pmc": null,
"reference": "Wang XJ, Song ZG, Jiao HC and Lin H. Dexamethasone facilitates lipid accumulation in chicken skeletal muscle. Stress 2012; 15: 443-456.",
"DOI": null,
"article": 105
},
{
"id": 3238,
"serial_number": 13,
"pmc": null,
"reference": "Sato AY., Richardson D, Cregor M, Davis HM, ED, Mcandrews K, Zimmers TA, Organ JM, Peacock M, Plotkin LI and Bellido T. Glucocorticoids Induce Bone and Muscle Atrophy by Tissue-Specific Mechanisms Upstream of E3 Ubiquitin Ligases. Endocrinol. 2017; 158: 664–677.",
"DOI": null,
"article": 105
},
{
"id": 3239,
"serial_number": 14,
"pmc": null,
"reference": "Fappi A, Neves JC, Sanches LN, Silva PVM, Sikusawa GY, Brandão TPC, Chadi G, and Zanoteli E. Skeletal Muscle Response to Deflazacort, Dexamethasone and Methylprednisolone. Cells 2019; 8: 406. 10.3390/cells8050406.",
"DOI": null,
"article": 105
},
{
"id": 3240,
"serial_number": 15,
"pmc": null,
"reference": "Yano AA, Iriyanti N, and Rosidi. Effect of supplemental fermeherbafit on total blood lipid and cholesterol in broiler chickens. J Phys 2018; 1025: 012057.",
"DOI": null,
"article": 105
},
{
"id": 3241,
"serial_number": 16,
"pmc": null,
"reference": "Mir NA, Rafiq A, Kumar F, Singh V, and Shukla V. Determinants of broiler chicken meat quality and factors affecting them: a review. Int. J Food Sci and Tech 2017; 54: 2997–3009.",
"DOI": null,
"article": 105
},
{
"id": 3242,
"serial_number": 17,
"pmc": null,
"reference": "Wideman N, O’bryan CA and Crandall PG. Factors affecting poultry meat colour and consumer preferences – A review. World Poultry Sci J 2016, 72: 354-366.",
"DOI": null,
"article": 105
},
{
"id": 3243,
"serial_number": 18,
"pmc": null,
"reference": "Mobini B. Histological properties of intramuscular connective tissues in native chickens and their relationship with meat tenderness. Anim Sci J 2015; 2: 105-109.",
"DOI": null,
"article": 105
},
{
"id": 3244,
"serial_number": 19,
"pmc": null,
"reference": "Ademu LA, Erakpatobor-Iyeghe GT, Barje PP, Daudu OM and Wafar RJ. Response of Broiler Chickens under Dexamethasone Induced Stress Conditions. AJAVA 2018; 1: 1-9.",
"DOI": null,
"article": 105
},
{
"id": 3245,
"serial_number": 20,
"pmc": null,
"reference": "Hussein MR, Hamed SA , Mostafa MG, Abu-Dief_ Nageh EE Kamel F and Kandil MR . The effects of glucocorticoid therapy on the inflammatory and Dendritic cells in muscular dystrophies. Int. J. Exp. Path. 2006, 87, 451–461.",
"DOI": null,
"article": 105
},
{
"id": 3246,
"serial_number": 21,
"pmc": null,
"reference": "Otrocka-Domagała I, Paździor-Czapula K and Gesek M. Dexamethasone-induced impairment of post-injury skeletal muscle regeneration. BMC Veterinary Research 2019.15:56.",
"DOI": null,
"article": 105
},
{
"id": 3247,
"serial_number": 22,
"pmc": null,
"reference": "Aengwanich W. Effects of dexamethasone on physiological changes and productive performance in broilers. AJAVA 2007; 32: 157-161.",
"DOI": null,
"article": 105
},
{
"id": 3248,
"serial_number": 23,
"pmc": null,
"reference": "Darda U, Warsch BS and Pereira D. High‐dose glucocorticoids improve renal failure reversibility in patients with newly diagnosed multiple myeloma. American Journal of Hematology, 2011 86: 224-227.",
"DOI": null,
"article": 105
},
{
"id": 3249,
"serial_number": 24,
"pmc": null,
"reference": "Eken H, Ozturk H, Ozturk H and Buyukbayram H. Dose-related effects of dexamethasone on liver damage due to bile duct ligation in rats. World J Gastroenterol 2006, 33: 5379-5383.Schakman O, Kalista S, Barbé C, Loumaye A and Thissen JP. Glucocorticoid-induced skeletal muscle atrophy. Int J Biochem Cell B. 2013; 45: 2163–2172.",
"DOI": null,
"article": 105
},
{
"id": 3250,
"serial_number": 25,
"pmc": null,
"reference": "Ramasamy K, Norhani A, Syed J, Michael W and Yin WHO. Effects of Lactobacillus feed supplementation on cholesterol, fat content and fatty acid composition of the liver, muscle and carcass of broiler chickens. Anim Res 2006; 55: 77–82.",
"DOI": null,
"article": 105
},
{
"id": 3251,
"serial_number": 26,
"pmc": null,
"reference": "Al-Ruwaili M, Herzallah S, Al-Dmoor H and Al-Atiyat R. Effect of Broiler Commercial Strains on Total and Free Cholesterol Levels of Chicken Muscle Tissues. Glob Vet 2014; 12: 381-383.",
"DOI": null,
"article": 105
}
]
},
{
"id": 99,
"slug": "178-1593105436-identification-of-potential-sars-cov-2-main-protease-inhibitors-from-ficus-carica-latex-an-in-silico-approach",
"featured": false,
"slider": false,
"issue": "Special Issue",
"type": "original_article",
"manuscript_id": "178-1593105436",
"recieved": "2020-06-25",
"revised": null,
"accepted": "2020-08-26",
"published": "2020-09-09",
"pdf_file": "https://jabet.bsmiab.org/media/pdf_file/2023/13/178-1593105436.pdf",
"title": "Identification of potential SARS-CoV-2 main protease inhibitors from Ficus Carica Latex: An in-silico approach",
"abstract": "<p>SARS-CoV-2 (severe acute respiratory syndrome coronavirus 2) is the aetiological agent behind the current pandemic of coronavirus disease 2019 (COVID-19). SARS-CoV-2 main protease plays a dynamic role in mediating viral replication and transcription, which is one of the most probable drug targets against SARS-CoV-2. <em>Ficus carica </em>latex encompasses notable bioactive molecules with various biological properties, including antiviral activities. In this study, latex compounds of <em>Ficus carica</em> were screened to find out active phytochemicals against SARS-CoV-2 main protease through molecular docking, molecular dynamics simulation, and ADMET (absorption, distribution, metabolism, excretion, and toxicity) profiling. A total of 21 compounds were screened, and the compounds, lupeol, α-amyrin, and luteolin, showed the highest binding affinity and intense interaction with the vital catalytic residue His 41 and Cys 145. The molecular dynamics simulation revealed that the amyrin is the most stable compound with higher binding free energy, suggesting that this compound can compete with the native ligands of the main protease. The ADMET analysis indicated that these phytochemicals have considerable physicochemical, pharmacokinetics, and drug-likeness properties and do not possess any considerable detrimental effects and can be considered as potential drug candidates against SARS-CoV-2. However, further in-vitro, in-vivo, and clinical trials are required to observe the exact efficiency of these compounds.</p>",
"journal_reference": "J Adv Biotechnol Exp Ther. 2020; 3(4): 57-67.",
"academic_editor": "Md. Masudur Rahman, PhD; Sylhet Agricultural University, Sylhet 3100, Bangladesh",
"cite_info": "Ali MC, Nur AJ, et al. Identification of potential SARS-CoV-2 main protease inhibitors from Ficus Carica Latex: An in-silico approach. J Adv Biotechnol Exp Ther. 2020; 3(4): 57-67.",
"keywords": [
"Molecular docking",
"SARS-CoV-2",
"COVID-19",
"Antiviral drugs",
"Ficus carica latex",
"Molecular dynamics"
],
"DOI": "10.5455/jabet.2020.d157",
"sections": [
{
"section_number": 1,
"section_title": "INTRODUCTION",
"body": "<p>SARS-CoV-2 is like an unexpected nightmare of 2020 that hampered the normal activities of humans and changed the way of life on earth. SARS-CoV-2 is a single-stranded, positive-sense, enveloped virus with a large RNA genome about ~30kb [<a href=\"#r-1\">1, 2</a>]. Unfortunately, there are no specific vaccines or drugs available to treat SARS-CoV-2, although various clinical trials are started to test the feasibility of some drugs. Meanwhile, several existing drugs are repurposed like remdesivir, chloroquine, hydroxychloroquine, lopinavir, and ritonavir against SARS-CoV-2 infection [<a href=\"#r-3\">3, 4</a>]. However, they have primarily proven effective, but still, their efficacy and safety are questionable [<a href=\"#r-5\">5-7</a>]. For these limitations, it is essential to find out the specific drug that would effectively inhibit or reduce SARS-CoV-2 infection.<br />\r\nThe protease enzyme of SARS-CoV-2 cleaves its native polypeptides and generates active fragments responsible for viral replication, transcription, and translation [<a href=\"#r-8\">8</a>]. These polypeptides are essential for viral replication, transcription, and translation [<a href=\"#r-9\">9, 10</a>]. The main protease (M<sup>pro</sup>) catalyzes the SARS-CoV-2 translated polyproteins’ relevance to the viral life cycle and makes them functional [<a href=\"#r-11\">11</a>]. “M<sup>pro </sup>has three functional domains; domain I (residues 8–101), domain II (residues 102–184), and domain III (residues 201–303) [<a href=\"#r-12\">12</a>].” “Domain II is joined with domain III by a loop (residues 185–200). The ligand attachment site is situated in the loop of domain I and domain II where catalytic dyad Cys 145-His 41 plays vital roles in ligand management [<a href=\"#r-12\">12</a>]” (shown in <a href=\"#figure1\">figure 1</a>). M<sup>pro</sup> also plays a significant role in SARS-CoV-2 replication [<a href=\"#r-12\">12</a>]. Besides, M<sup>pro </sup>is not functionally correlated with the human’s homologous proteases, which implies M<sup>pro </sup>as a striking target for drug designing [<a href=\"#r-12\">12</a>]. Moreover, several previous studies have shown that M<sup>pro </sup>is the prominent target against SARS-CoV-2 infection [<a href=\"#r-13\">13-15</a>]. However, nature provides the best remedy. In previous studies, it has been shown that natural phytochemicals contain active ingredients that might be able to inhibit the SARS-CoV-2 M<sup>pro </sup> [<a href=\"#r-14\">14</a>, <a href=\"#r-16\">16, 17</a>].<br />\r\nThe <em>Ficus carica</em> latex (F-latex) encompasses notable molecules with remarkable benefits against antimicrobial-resistant microorganisms along with other biological properties [<a href=\"#r-18\">18, 19</a>]. F-latex contains fatty acids, flavonoids, phenolics, tannins, alkaloids, terpenoids, and sterols with numerous biological benefits [<a href=\"#r-19\">19, 20</a>]. It has been used as traditional medicine around the world and found no cytotoxicity on Vero cells [<a href=\"#r-21\">21</a>]. Several studies found that the F-latex extracts have antibacterial [<a href=\"#r-22\">22, 23</a>], antifungal [<a href=\"#r-22\">22</a>], and antiviral [<a href=\"#r-21\">21</a>] properties. The F-latex compound is active against both DNA and RNA viruses including, adenovirus (ADV), echovirus type 11 (ECV-11) [<a href=\"#r-21\">21</a>], herpes simplex type 1 (HSV-1) [<a href=\"#r-21\">21</a>], HSV-2 [<a href=\"#r-24\">24</a>], caprine herpesvirus-1 (CpHV-1) (showed almost similar efficiency like acyclovir) [<a href=\"#r-25\">25</a>], human papillomavirus (HPV) [26], and the influenza virus H9N2 [<a href=\"#r-27\">27</a>]. As previous studies showed that the F-latex compounds have potential antiviral activity against several viruses along with the same realm, riboviria, so it might be possible to act against SARS-CoV-2 [<a href=\"#r-27\">27</a>]. Therefore, the present study was intended to discover the SARS-CoV-2 M<sup>pro</sup> inhibitors from the F-latex compounds to fight against SARS-CoV-2 infection.</p>\r\n\r\n<div id=\"figure1\">\r\n<figure class=\"image\"><img alt=\"\" height=\"262\" src=\"/media/article_images/2024/36/07/178-1593105436-Figure1.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 1. </strong>The 2D structure of main protease. Here, the schematic diagram of domain I is represented by yellow color, domain II is represented by blue-green color, domain III is represented by dark red color, and the catalytic Cys-His dyad is represented in CYK form by red color.</figcaption>\r\n</figure>\r\n</div>"
},
{
"section_number": 2,
"section_title": "MATERIALS AND METHODS",
"body": "<p><strong><em>Ficus carica</em></strong><strong> latex compounds profiling and ligand preparation</strong><br />\r\nIn this study, we consider <em>Ficus carica</em> latex as a reservoir of SARS-CoV-2 inhibitory compounds. We prepared a dataset of the active phytochemicals of <em>Ficus carica </em>latex from prior studies and the findings. We made a library of bioactive latex compounds by searching related literature in PubMed, google scholar, the web of science, and Scopus databases [<a href=\"#r-28\">28</a>]. We used the PubChem database [<a href=\"#r-29\">29</a>] to download the SDF format of the 3D structures of all compounds and used Open Babel software to convert SDF format compounds to PDB format [<a href=\"#r-30\">30</a>]. We used PyRx [<a href=\"#r-31\">31</a>] integrated mmff94 (Merck molecular force field<strong>) </strong>force field [<a href=\"#r-32\">32</a>] for optimization and ligand preparation. For further analysis, the ligands were then converted into PDBQT format.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Preparation of receptor</strong><br />\r\nThe crystallized structure of M<sup>pro </sup>(PDB ID 6LU7 [<a href=\"#r-12\">12</a>]) was curated from the RCSB PDB resource [<a href=\"#r-33\">33</a>]. The water molecules and ligand were removed from the receptor and energy minimized by steepest descent and conjugate gradient techniques. The GROMACS 96 43B1 algorithm in SWISS-PDB viewer [<a href=\"#r-34\">34</a>], and Chimera (Amber Force field) were utilized to prepare the final receptor [<a href=\"#r-35\">35</a>].</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Virtual screening</strong><br />\r\nVirtual screening of all compounds was executed through PyRx software by AutoDock wizard [<a href=\"#r-31\">31</a>, <a href=\"#r-36\">36</a>]. Ligands were considered as flexible, and the protein was considered as rigid during docking. The auto grid engine in AutoDock was used to generate the configuration file of grid parameters (grid box size X, Y, Z; 23, 28, 29, respectively). Potential compounds were considered based on RMSD (root mean square deviation) values. Ligands with most negative docking scores and lower (<1Å) RMSD values were considered for further investigation. The molecular interactions between ligands and receptors were visualized using the Biovia discovery studio visualizer (v 4.5) [<a href=\"#r-37\">37</a>].</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Molecular dynamics simulation</strong><br />\r\nIn order to get more insights of the protein-ligand complex, molecular dynamics simulation (MD simulation) was performed. The selected complexes were subjected to MD simulation by YASARA molecular dynamics software package using AMBER1 14 forcefield [<a href=\"#r-38\">38, 39</a>] as described formerly [<a href=\"#r-40\">40-42</a>]. The complex was solvated inside the simulation cell using the TIP3 water model [<a href=\"#r-43\">43</a>]. The periodic boundary condition was optimized, and water and Na<sup>+</sup>/Cl<sup>– </sup>ions were added. The steepest gradient approach (5000 cycles) was used for each simulation system to minimize energy. We used the PME method to explain the long-range electrostatic interactions within the following conditions 8 Å (electrostatic interaction cut off distance), 298 K temp., 0.9% NaCl, and pH of 7.4 [<a href=\"#r-44\">44</a>]. The simulation time step interval was set to 2 fs [<a href=\"#r-45\">45</a>]. Finally, the simulation was conducted for 20 ns in Berendsen thermostat [<a href=\"#r-46\">46</a>], where the snapshots were saved for every 10 ps. The VMD [<a href=\"#r-47\">47, 48</a>] software was used to calculate RMSD (root mean square deviation), RMSF (root mean square fluctuations), Rg (radius of gyration), and SASA (solvent accessible surface area). Furthermore, the Molecular Mechanics-Poisson–Boltzmann Surface Area (MM-PBSA) binding free energy calculation method was used for all systems to calculate the binding free energy by the following formula,<br />\r\nBinding Energy = E<sub>potRecept</sub> + E<sub>solvRecept</sub> + E<sub>potLigand</sub> + E<sub>solvLigand</sub> – E<sub>potComplex</sub> – E<sub>solvComplex</sub></p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>ADMET analysis</strong><br />\r\nThe compounds that have a higher binding affinity (-12.5 to -7.4) were considered for further ADMET analysis. Based on the canonical SMILES of chosen ligands obtained from PubChem, ADME properties of potent drug candidates were calculated by SwissADME [49], whereas toxicity was analyzed using PreADMET toxicity prediction tools [<a href=\"#r-50\">50</a>].</p>"
},
{
"section_number": 3,
"section_title": "RESULTS",
"body": "<p><strong>Virtual screening of retrieved compounds against SARS-CoV-2 main protease</strong><br />\r\nVirtual screening is an effective method to find out probable hits from thousands of compounds [<a href=\"#r-51\">51</a>]. Virtual screening utilizing molecular docking is an alternative way for lead identification in drug discovery [<a href=\"#r-51\">51, 52</a>]. We conducted molecular docking for all (total 21) identified compounds (<a href=\"#Table-1\">table 1</a>) with SARS-CoV-2 main protease. The binding affinity of the compounds lies in the following ranges -12.5 to -10.2 kcal/mol, -10.2 to -7.9 kcal/mol, -7.9 to -5.6 kcal/mol, and -5.6 to -5.3 kcal/mol (<a href=\"#figure2\">figure 2</a>). According to the binding affinity, three compounds were selected, namely lupeol (-12.5 kcal/mol), α-amyrin (-7.9 kcal/mol), and luteolin (-7.4 kcal/mol), and considered for further analysis. The a-ketoamide was used as a control ligand in this study as it has been identified recently as a potent inhibitor (binding affinity -7.3 kcal/mol) against SARS-CoV-2 main protease [<a href=\"#r-53\">53</a>]. The α-ketoamide and selected compounds binding affinity, Pubchem Id, formula, IUPAC name, and 2D structure are depicted in <a href=\"#Table-2\">table 2</a>.</p>\r\n\r\n<div id=\"figure2\">\r\n<figure class=\"image\"><img alt=\"\" height=\"354\" src=\"/media/article_images/2024/36/07/178-1593105436-Figure2.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 2.</strong> Docking scores of all compounds. Most of the compounds have moderate binding affinity.</figcaption>\r\n</figure>\r\n</div>\r\n\r\n<div id=\"Table-1\">\r\n<p><a href=\"https://jabet.bsmiab.org/table/178-1593105436-table1/\">Table-1</a><strong>Table 1. </strong>Binding affinity of latex compounds of <em>Ficus carica</em> latex.</p>\r\n</div>\r\n\r\n<div id=\"Table-2\">\r\n<p><a href=\"https://jabet.bsmiab.org/table/178-1593105436-table2/\">Table-2</a><strong>Table 2. </strong>Binding affinity, PubChem Id, formula, binding affinity, IUPAC name, and 2D structure of selected SARS-CoV-2 M<sup>pro</sup> inhibitors.</p>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Molecular interaction analysis of selected compounds</strong><br />\r\nThe molecular interactions of the selected compounds were visualized using BIOVIA discovery studio visualizer (v 4.5) [<a href=\"#r-37\">37</a>]. In a recent study, Zhang et al. showed that the inhibitor a-ketoamide forms molecular interactions with His 41, Gly 143, Ser 144, Cys 145, His 163, His 164, Glu 166, Pro 168, and Gln 189 residue of the main protease [<a href=\"#r-53\">53</a>]. Moreover, the native ligand (N3) of our selected M<sup>pro</sup> (6LU7) also interacts with the catalytic dyad Cys 145-His 41 [<a href=\"#r-11\">11</a>]. The docked compounds interaction indicates that all three compounds interact with either catalytic residues His 41 and Cys 145 or at least one of them. Moreover, the ligand forms interaction with other substrate-binding pocket residues shown in<a href=\"#figure3\"> figure 3</a>. The positive control, a-ketoamide, formed three strong hydrogen bonds with the catalytic loops’ residue Gln 189 and several alkyl bonds with residues Leu 27, His 41, Met 49, Cys145, and Met 165 (<a href=\"#figure3\">figure 3b</a>). Lupeol forms alkyl bonds with Leu 27, His 41, Met 49, and Cys 145 residue (<a href=\"#figure3\">figure 3c</a>). Amyrin forms several alkyl bonds with His 41, Met 49, Cys 145, and Met 165 residue (<a href=\"#figure3\">figure 3d</a>). Luteolin forms hydrogen bonds with Cys 145 and Thr 190 and alkyl bonds with Met 165 residue (<a href=\"#figure23\">figure 3e</a>).</p>\r\n\r\n<div id=\"figure3\">\r\n<figure class=\"image\"><img alt=\"\" height=\"306\" src=\"/media/article_images/2024/36/07/178-1593105436-Figure3.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 3.</strong> Molecular interactions of selected compounds with SARS-Cov-2 main protease (Mpro). (a) Molecular interactions of all latex compounds, the interaction of (b) positive control α-ketoamide with Mpro, (c) lupeol-Mpro, (d) α-amyrin-Mpro, and (e) luteolin- Mpro. All selected compounds interact with the vital substrate management catalytic dyad Cys 145-His 41.</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Molecular dynamics simulation</strong><br />\r\nThe protein-ligand complex stability was analyzed through MD simulation. We used four parameters to understand complex stability, including RMSD, RMSF, Rg, and SASA. In <a href=\"#figure4\">figure 4a</a>, the positive control α-ketoamide showed the lowest RMSD values compared to all systems, including the apo-proteins. The lower RMSD value illustrates the compactness of the system. The amyrin-protein complex showed a quite similar fashion of stability like apo-protein with stable conformation at 3 ns to 8 ns, and 12 ns to 20 ns. The lupeol also showed stable conformation at 2 ns to 11 ns; after that, a significant structural drifting occurs around 11.5 ns later on, again showed stability from 12 ns to the rest of the simulation. The luteolin complex showed stability at 5 ns to 10 ns, and 18 ns to 20 ns, whereas the rest of the time, it showed more fluctuation. The average RMSD values were 1.43 Å, 1.28 Å, 1.51 Å, 1.66 Å, and 1.98 Å for apo-protein, α-ketoamide-M<sup>pro</sup>, amyrin-M<sup>pro</sup>, lupeol- M<sup>pro</sup>, and luteolin-M<sup>pro</sup> complex, respectively (shown in<a href=\"#Table-3\"> table 3</a>).<br />\r\nIn addition, the RMSF calculation was used to analyze the local residual change (shown in figure 4b). The higher RMSF values illustrate the higher flexibility of the system. The α-ketoamide-M<sup>pro</sup> complex showed a lower average RMSF value (1.14 Å) compared to all systems. The amyrin- M<sup>pro </sup>average RMSF value (1.21 Å) was quite similar to apo-protein RMSF value 1.22 Å. However, lupeol-M<sup>pro</sup> and luteolin-M<sup>pro</sup> showed 1.35 Å and 1.28 Å RMSF values, respectively (shown in <a href=\"#Table-3\">table 3</a>). Besides, the structural compactness was analyzed by calculating the radius of gyration (figure 4c). The lower RG values represent the compactness of the system. Here, the positive control α-ketoamide-M<sup>pro</sup> complex showed lower Rg value (22.37 Å) compared to the apo-protein (23.35 Å), while, the amyrin-M<sup>pro </sup>complex showed almost same Rg value (22.40 Å) like the positive control. However, the lupeol-M<sup>pro</sup> and luteolin-M<sup>pro</sup> complex showed lower Rg values, 22.47 Å, and 22.31 Å, respectively, than apo-protein (shown in <a href=\"#Table-3\">table 3</a>).<br />\r\nMoreover, the solvent-accessible surface area (SASA) of all systems were analyzed and depicted in figure 4d. The lower SASA value represents the compactness of the systems. The average SASA value of the amyrin-M<sup>pro </sup>complex was lower (14059.42 nm<sup>2</sup>) than the positive control α-ketoamide-M<sup>pro</sup> complex (14171.00 nm<sup>2</sup>). The SASA values of the lupeol-M<sup>pro</sup> and luteolin-M<sup>pro</sup> complex were 14248.64 nm<sup>2</sup> and 14263.55 nm<sup>2</sup>, respectively (shown in <a href=\"#Table-3\">table 3</a>).<br />\r\nFurthermore, the binding free energy calculation depicted that the amyrin showed higher binding energy (225.896 kJ/mol), whereas the lupeol and luteolin showed 181.434 kJ/mol and 51.063 kJ/mol binding free energy, respectively (shown in <a href=\"#Table-3\">table 3</a>).</p>\r\n\r\n<div id=\"figure4\">\r\n<figure class=\"image\"><img alt=\"\" height=\"281\" src=\"/media/article_images/2024/36/07/178-1593105436-Figure4.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 4.</strong> Molecular dynamics simulation. Analysis of (a) RMSD (Root Mean Square Deviation); (b) RMSF (Root Mean Square Fluctuations); (c) Rg (Radius of Gyration); (d) SASA (Solvent Accessible Surface Area); and (e) Binding free energy.</figcaption>\r\n</figure>\r\n</div>\r\n\r\n<div id=\"Table-3\">\r\n<p><a href=\"https://jabet.bsmiab.org/table/178-1593105436-table3/\">Table-3</a><strong>Table 3. </strong>The average mean value of MD trajectory.</p>\r\n</div>\r\n\r\n<div id=\"Table-4\">\r\n<p><a href=\"https://jabet.bsmiab.org/table/178-1593105436-table4/\">Table-4</a><strong>Table 4. </strong>ADMET analysis of top 3 compounds.</p>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>ADMET analysis</strong><br />\r\nThe different ADMET properties of the selected three compounds are depicted in <a href=\"#Table-4\">table 4</a>. In this study, we analyzed physicochemical properties, lipophilicity, water-solubility, pharmacokinetics, drug-likeness, medicinal chemistry, and toxic properties of selected compounds employing ADMET analysis. The pharmacokinetics properties show that the selected compounds cannot permeate the blood-brain barrier (BBB), do not inhibits any cytochrome P<sup>450</sup> isoforms (lupeol and α- amyrin), and luteolin shows higher GI absorption, in contrast, lupeol and α-amyrin shows lower GI absorption. In terms of drug-likeness activity, luteolin does not violate Lipinski’s role of five, but both lupeol and α-amyrin violate one role. The toxicity analysis result shows that these compounds do not inhibit the human ether go-go gene (hERG). The hERG gene maintains cardiac systolic and diastolic activity through the potassium ion channel, and inhibition of these channels disrupts the homeostatic balance. The Ames test toxicity data (in TA100 strain-metabolic activation by rat liver homogenate) also shows that these compounds are not mutagenic. However, the detailed ADMET interpretation is explained in the discussion section.</p>"
},
{
"section_number": 4,
"section_title": "DISCUSSION",
"body": "<p>Although some drugs are repurposed, there is an urgent need for effective and specific drugs against SARS-CoV-2 infection cause the repurposed drugs have been shown some drastic side effects [<a href=\"#r-54\">54, 55</a>]. Structure-based virtual screening (VS) has been a trend in drug development with all of its underlying computational approaches for more than a decade now, and molecular docking has been thoroughly studied [<a href=\"#r-51\">51</a>]. In this study, we used computer-aided techniques to identify potential inhibitors of SARS-CoV-2 M<sup>pro</sup>. The F-latex extracts have been found to inhibit the replication of HSV-1, ECV-11, CpHV-1, and ADV [<a href=\"#r-21\">21</a>, <a href=\"#r-25\">25</a>]. Moreover., they have been found to act against several drug-resistant pathogens [<a href=\"#r-56\">56</a>].<br />\r\nHowever, among the 21 studied F-latex compounds, the top three compounds are selected considering their highest binding affinity, and it is found that lupeol, α-amyrin, and luteolin could be used as promising inhibitors against SARS-CoV-2 main protease. According to the binding affinity, lupeol (-12.5 kcal/mol), α-amyrin (-7.9 kcal/mol), and luteolin (-7.4 kcal/mol) have a higher binding affinity than the control α-ketoamide (-7.3 kcal/mol) shown in <a href=\"#Table-2\">table 2</a>. Lupeol forms non-covalent alkyl bonds with the crucial catalytic residue His 41 and Cys 145 like α-amyrin. In contrast, luteolin forms one hydrogen and alkyl bonds with Cys 145. Thereby, it may act as inhibitors of SARS-CoV-2 main protease. The selected three compounds form strong non-covalent interactions with other binding site residues. However, similar recent studies also support these findings where the inhibitor compounds form strong covalent and non-covalent bonds with the following residues His 41, Met 49, Tyr 54, Phe 140, His 164, Met 165, Glu 166, Pro168, Asp 187, Arg 188, and Gln189 [<a href=\"#r-57\">57, 58</a>]. Some recent studies suggest that lupeol has antiviral activity against hepatitis B [<a href=\"#r-59\">59</a>], HIV [<a href=\"#r-60\">60</a>], and also reduces pro-inflammatory cytokines [<a href=\"#r-61\">61</a>]. It is also active as an antioxidant, anti-dyslipidemia, anti-hyperglycemic, and anti-mutagenic agent. Besides, it shows hepatoprotective, nephroprotective, neuroprotective, cardioprotective, and anticancer activity in in-vitro and in vivo experiments [<a href=\"#r-62\">62</a>]. Amyrin also has antiviral, anti-inflammatory, and antioxidants properties [<a href=\"#r-59\">59</a>]. The flavonoid luteolin is a potential antiviral agent that is validated by several experiments’ models like as it shows antiviral activity against the tick-borne encephalitis virus model [<a href=\"#r-63\">63</a>], Japanese encephalitis virus [<a href=\"#r-64\">64</a>], HIV-1 [<a href=\"#r-65\">65</a>], and Coxsackie virus B3 [<a href=\"#r-66\">66</a>].<br />\r\nMolecular dynamics simulation acts as a computational microscope to provide bio-molecular insights at progressive and conformational scales [<a href=\"#r-67\">67</a>]. The higher RMSD, RMSF, Rg, and SASA values denote higher flexibility of the system [<a href=\"#r-68\">68, 69</a>]. The α-ketoamide showed lower RMSD, RMSF, and Rg values, which represent its stable binding with SARS-CoV-2 main protease. The higher SASA value of α-ketoamide depicts the system conformation drifting. The amyrin had lesser RMSF, SASA, and Rg values compared to the apo-protein, which depicts its conformational compactness. Besides, the highest binding free energy represents its better binding ability with SARS-CoV-2 main protease. The lupeol showed lower Rg value in contrast with apo-protein and higher binding energy compared to the α-ketoamide, which represents a strong binding, and flexible conformation of this system. Higher RMSD, RMSF, Rg, and SASA values of luteolin represented its higher flexibility compared to all systems. The lupeol and luteolin were the least compacts compounds according to the analysis.<br />\r\nIn silico ADMET analysis is a productive, comprehensive, timely, and cost-saving approach to analyze the physicochemical, and drug-likeness properties of any compounds [<a href=\"#r-70\">70</a>]. Computational biology contribution has speeded up drug discovery and is being used to find and develop novel lead compounds against many pathogenic microorganisms and diseases in the biopharmaceutical sector [<a href=\"#r-71\">71, 72</a>]. ADMET analysis provides a clear image of possible drug candidates. The optimal molecular weight of a possible drug should be between 150 to 500 g/mol (Dalton), hydrogen bond acceptor (HBA) ≤ 10, hydrogen bond donor (HBD) ≤ 5, TPSA (topological polar surface area) between 20 and 130 Å<sup>2</sup>, and molar refractory (MR) range between 40 to 130 [<a href=\"#r-28\">28</a>, <a href=\"#r-49\">49</a>, <a href=\"#r-73\">73</a>]. In terms of physicochemical properties, our three selected compounds are in the acceptable range (<a href=\"#Table-4\">table 4</a>). The water solubility analysis reveals that luteolin is soluble in water, and the rests two are poorly soluble.<br />\r\nMoreover, these compounds do not penetrate BBB. In new drug discovery approaches, finding out the interaction of drug molecules with cytochrome P<sup>450</sup> (CYP) is important cause CYP isoforms maintains the normal cellular metabolism, transformation, and excretion of drugs. Our studied compounds lupeol and α-amyrin do not inhibit any CYP isoforms, which is another good sign of their effectiveness. The medicinal chemistry properties of lupeol and α-amyrin compounds show that they do not possess any PAINS (pan assay interference compounds) alert means they have a high tendency to bind specifically with their targets, and they do not exert any false-positive results. However, lupeol and α-amyrin violate lead likeness properties, but luteolin does not violate lead likeness properties. The studied compounds have moderate synthetic accessibility (value 1 for low and 10 for high synthetic accessibility) [<a href=\"#r-74\">74</a>].</p>\r\n\r\n<div id=\"figure5\">\r\n<figure class=\"image\"><img alt=\"\" height=\"371\" src=\"/media/article_images/2024/36/07/178-1593105436-Figure5.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 5. </strong>Schematic diagram of the summary. </figcaption>\r\n</figure>\r\n</div>"
},
{
"section_number": 5,
"section_title": "CONCLUSION",
"body": "<p>In sum, the virtual screening revealed that lupeol, α-amyrin, and luteolin show the highest binding affinity with SARS-CoV-2 main protease and show significant and vital interaction with either both catalytic residues His 41 and Cys 145 or at least one of them. The selected compounds also represent other meaningful interactions like hydrogen bonds, van der Waals bonds, and alkyl bonds. The molecular dynamics simulation study illustrated that among all the phytochemicals, amyrin is the most stable phytochemicals, which could be able to inhibit SARS-CoV-2 main protease strongly. The binding free energy analysis revealed that amyrin and lupeol have higher binding free energy compared to the known SARS-CoV-2 M<sup>pro</sup> inhibitor α-ketoamide. The drug-likeness efficiency of these compounds was analyzed by ADMET and reveled that they follow Lipinski’s rule of five and Veber rules with low or no toxicity. Moreover, these compounds cannot penetrate BBB, and in sum, they can be used as drug candidates against SARS-CoV-2.</p>"
},
{
"section_number": 6,
"section_title": "AUTHORS CONTRIBUTION",
"body": "<p>RD and MMR conceived the plan of this research. MCA, AJN, MSK wrote the manuscript. MCA analyzed the data and made the figures. RD and MMR, MMK edited the manuscript. All authors revised and approved the manuscript for final submission.</p>"
},
{
"section_number": 7,
"section_title": "ACKNOWLEDGEMENT",
"body": "<p>We are thankful to Yeasmin Akter Munni, Graduate Student, Department of Anatomy, Dongguk University College of Medicine, Gyeongju 38066, Republic of Korea, for helping to draw the figures of this project.</p>"
},
{
"section_number": 8,
"section_title": "CONFLICTS OF INTEREST",
"body": "<p>The authors declared no competing interests.</p>"
}
],
"figures": [
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/36/07/178-1593105436-Figure1.jpg",
"caption": "Figure 1. The 2D structure of main protease. Here, the schematic diagram of domain I is represented by yellow color, domain II is represented by blue-green color, domain III is represented by dark red color, and the catalytic Cys-His dyad is represented in CYK form by red color.",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/36/07/178-1593105436-Figure2.jpg",
"caption": "Figure 2. Docking scores of all compounds. Most of the compounds have moderate binding affinity.",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/36/07/178-1593105436-Figure3.jpg",
"caption": "Figure 3. Molecular interactions of selected compounds with SARS-Cov-2 main protease (Mpro). (a) Molecular interactions of all latex compounds, the interaction of (b) positive control α-ketoamide with Mpro, (c) lupeol-Mpro, (d) α-amyrin-Mpro, and (e) luteolin- Mpro. All selected compounds interact with the vital substrate management catalytic dyad Cys 145-His 41.",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/36/07/178-1593105436-Figure4.jpg",
"caption": "Figure 4. Molecular dynamics simulation. Analysis of (a) RMSD (Root Mean Square Deviation); (b) RMSF (Root Mean Square Fluctuations); (c) Rg (Radius of Gyration); (d) SASA (Solvent Accessible Surface Area); and (e) Binding free energy.",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/36/07/178-1593105436-Figure5.jpg",
"caption": "Figure 5. Schematic diagram of summary.",
"featured": false
}
],
"authors": [
{
"id": 378,
"affiliation": [
{
"affiliation": "Department of Biotechnology and Genetic Engineering, Faculty of Biological Sciences, Islamic University, Kushtia-7003, Bangladesh"
}
],
"first_name": "Md. Chayan",
"family_name": "Ali",
"email": null,
"author_order": 1,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 99
},
{
"id": 379,
"affiliation": [
{
"affiliation": "Department of Biotechnology and Genetic Engineering, Faculty of Biological Sciences, Islamic University, Kushtia-7003, Bangladesh"
}
],
"first_name": "Anjumana Jannati",
"family_name": "Nur",
"email": null,
"author_order": 2,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 99
},
{
"id": 380,
"affiliation": [
{
"affiliation": "Department of Biotechnology and Genetic Engineering, Faculty of Biological Sciences, Islamic University, Kushtia-7003, Bangladesh"
}
],
"first_name": "Mst. Shanzeda",
"family_name": "Khatun",
"email": null,
"author_order": 3,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 99
},
{
"id": 381,
"affiliation": [
{
"affiliation": "Department of Anatomy, Dongguk University College of Medicine, Gyeongju 38066, Republic of Korea."
}
],
"first_name": "Raju",
"family_name": "Dash",
"email": "rajudash.bgctub@gamil.com",
"author_order": 4,
"ORCID": null,
"corresponding": true,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "Raju Dash, Department of Anatomy, Dongguk University College of Medicine, Gyeongju 38066, Republic of Korea. Email: rajudash.bgctub@gamil.com",
"article": 99
},
{
"id": 382,
"affiliation": [
{
"affiliation": "Department of Biotechnology and Genetic Engineering, Faculty of Biological Sciences, Islamic University, Kushtia-7003, Bangladesh"
}
],
"first_name": "Md Mafizur",
"family_name": "Rahman",
"email": "shilon83@gmail.com",
"author_order": 5,
"ORCID": null,
"corresponding": true,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "Md Mafizur Rahman, Department of Biotechnology and Genetic Engineering, Faculty of Biological Sciences, Islamic University, Kushtia-7003, Bangladesh. Email: shilon83@gmail.com",
"article": 99
},
{
"id": 383,
"affiliation": [
{
"affiliation": "Department of Biotechnology and Genetic Engineering, Faculty of Biological Sciences, Islamic University, Kushtia-7003, Bangladesh"
}
],
"first_name": "Mohammad Minnatul",
"family_name": "Karim",
"email": null,
"author_order": 6,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 99
}
],
"views": 1260,
"downloads": 175,
"references": [
{
"id": 3070,
"serial_number": 1,
"pmc": null,
"reference": "Bosch BJ, van der Zee R, de Haan CA, Rottier PJ. The coronavirus spike protein is a class I virus fusion protein: structural and functional characterization of the fusion core complex. J Virol. 2003;77:8801-11.",
"DOI": null,
"article": 99
},
{
"id": 3071,
"serial_number": 2,
"pmc": null,
"reference": "Wu F, Zhao S, Yu B, Chen Y-M, Wang W, Hu Y, et al. Complete genome characterisation of a novel coronavirus associated with severe human respiratory disease in Wuhan, China. bioRxiv. 2020.",
"DOI": null,
"article": 99
},
{
"id": 3072,
"serial_number": 3,
"pmc": null,
"reference": "Wang M, Cao R, Zhang L, Yang X, Liu J, Xu M, et al. Remdesivir and chloroquine effectively inhibit the recently emerged novel coronavirus (2019-nCoV) in vitro. Cell Res. 2020;30:269-71.",
"DOI": null,
"article": 99
},
{
"id": 3073,
"serial_number": 4,
"pmc": null,
"reference": "Colson P, Rolain JM, Lagier JC, Brouqui P, Raoult D. Chloroquine and hydroxychloroquine as available weapons to fight COVID-19. Int J Antimicrob Agents. 2020;55:105932.",
"DOI": null,
"article": 99
},
{
"id": 3074,
"serial_number": 5,
"pmc": null,
"reference": "Magagnoli J, Narendran S, Pereira F, Cummings T, Hardin JW, Sutton SS, et al. Outcomes of hydroxychloroquine usage in United States veterans hospitalized with Covid-19. MedRxiv. 2020.",
"DOI": null,
"article": 99
},
{
"id": 3075,
"serial_number": 6,
"pmc": null,
"reference": "McGonagle D, O’Donnell JS, Sharif K, Emery P, Bridgewood C. Immune mechanisms of pulmonary intravascular coagulopathy in COVID-19 pneumonia. Lancet Rheumatol. 2020.",
"DOI": null,
"article": 99
},
{
"id": 3076,
"serial_number": 7,
"pmc": null,
"reference": "Mehra MR, Desai SS, Ruschitzka F, Patel AN. RETRACTED: Hydroxychloroquine or chloroquine with or without a macrolide for treatment of COVID-19: a multinational registry analysis. Lancet (London, England). 2020.",
"DOI": null,
"article": 99
},
{
"id": 3077,
"serial_number": 8,
"pmc": null,
"reference": "Ul Qamar MT, Alqahtani SM, Alamri MA, Chen LL. Structural basis of SARS-CoV-2 3CL(pro) and anti-COVID-19 drug discovery from medicinal plants. J Pharm Anal. 2020.",
"DOI": null,
"article": 99
},
{
"id": 3078,
"serial_number": 9,
"pmc": null,
"reference": "Xue X, Yu H, Yang H, Xue F, Wu Z, Shen W, et al. Structures of two coronavirus main proteases: implications for substrate binding and antiviral drug design. J Virol. 2008;82:2515-27.",
"DOI": null,
"article": 99
},
{
"id": 3079,
"serial_number": 10,
"pmc": null,
"reference": "Wang F, Chen C, Tan W, Yang K, Yang H. Structure of Main Protease from Human Coronavirus NL63: Insights for Wide Spectrum Anti-Coronavirus Drug Design. Sci Rep. 2016;6:22677.",
"DOI": null,
"article": 99
},
{
"id": 3080,
"serial_number": 11,
"pmc": null,
"reference": "Jin Z, Du X, Xu Y, Deng Y, Liu M, Zhao Y, et al. Structure of M(pro) from SARS-CoV-2 and discovery of its inhibitors. Nature. 2020;582:289-93.",
"DOI": null,
"article": 99
},
{
"id": 3081,
"serial_number": 12,
"pmc": null,
"reference": "Pillaiyar T, Manickam M, Namasivayam V, Hayashi Y, Jung SH. An Overview of Severe Acute Respiratory Syndrome-Coronavirus (SARS-CoV) 3CL Protease Inhibitors: Peptidomimetics and Small Molecule Chemotherapy. J Med Chem. 2016;59:6595-628.",
"DOI": null,
"article": 99
},
{
"id": 3082,
"serial_number": 13,
"pmc": null,
"reference": "Das S, Sarmah S, Lyndem S, Singha Roy A. An investigation into the identification of potential inhibitors of SARS-CoV-2 main protease using molecular docking study. J Biomol Struct Dyn. 2020:1-11.",
"DOI": null,
"article": 99
},
{
"id": 3083,
"serial_number": 14,
"pmc": null,
"reference": "Gentile D, Patamia V, Scala A, Sciortino MT, Piperno A, Rescifina A. Putative Inhibitors of SARS-CoV-2 Main Protease from A Library of Marine Natural Products: A Virtual Screening and Molecular Modeling Study. Mar Drugs. 2020;18.",
"DOI": null,
"article": 99
},
{
"id": 3084,
"serial_number": 15,
"pmc": null,
"reference": "Bacha U, Barrila J, Velazquez-Campoy A, Leavitt SA, Freire E. Identification of novel inhibitors of the SARS coronavirus main protease 3CLpro. Biochemistry. 2004;43:4906-12.",
"DOI": null,
"article": 99
},
{
"id": 3085,
"serial_number": 16,
"pmc": null,
"reference": "Cheng B, Li T. Discovery of alliin as a putative inhibitor of the main protease of SARS-CoV-2 by molecular docking. BioTechniques. 2020.",
"DOI": null,
"article": 99
},
{
"id": 3086,
"serial_number": 17,
"pmc": null,
"reference": "Ton AT, Gentile F, Hsing M, Ban F, Cherkasov A. Rapid Identification of Potential Inhibitors of SARS-CoV-2 Main Protease by Deep Docking of 1.3 Billion Compounds. Mol Inform. 2020.",
"DOI": null,
"article": 99
},
{
"id": 3087,
"serial_number": 18,
"pmc": null,
"reference": "Ripert C, Pierre A, Jean-Marie B, Anne-Pauline B, Claude G, Mycologie médicale: Tec & doc-Lavoisier; 2013.",
"DOI": null,
"article": 99
},
{
"id": 3088,
"serial_number": 19,
"pmc": null,
"reference": "Barolo MI, Ruiz Mostacero N, López SN. Ficus carica L. (Moraceae): an ancient source of food and health. Food Chem. 2014;164:119-27.",
"DOI": null,
"article": 99
},
{
"id": 3089,
"serial_number": 20,
"pmc": null,
"reference": "Al-Snafi AE. Nutritional and pharmacological importance of Ficus carica-A. J Pharm. 2017;7:22-48.",
"DOI": null,
"article": 99
},
{
"id": 3090,
"serial_number": 21,
"pmc": null,
"reference": "Lazreg Aref H, Gaaliche B, Fekih A, Mars M, Aouni M, Pierre Chaumon J, et al. In vitro cytotoxic and antiviral activities of Ficus carica latex extracts. Nat Prod Res. 2011;25:310-9.",
"DOI": null,
"article": 99
},
{
"id": 3091,
"serial_number": 22,
"pmc": null,
"reference": "Rashid KI, Mahdi NM, Alwan MA, Khalid LB. Antimicrobial activity of fig (Ficus carica Linn.) leaf extract as compared with latex extract against selected bacteria and fungi. J Univ Babylon. 2014;22:1620-6.",
"DOI": null,
"article": 99
},
{
"id": 3092,
"serial_number": 23,
"pmc": null,
"reference": "AL-Sabawi NA. The Antibacterial Effect of Fig (Leaves Extract and Latex) on Enterococcus faeca-lis as Intracanal Medicament.(An in vi-tro study). Al-Raf Den J. 2010;10:62-71.",
"DOI": null,
"article": 99
},
{
"id": 3093,
"serial_number": 24,
"pmc": null,
"reference": "Ay E, Duran N. Investigation of the antiviral activity of Ficus carica L. latex against HSV-2. International Conference on Advanced Materials and Systems (ICAMS): The National Research & Development Institute for Textiles and Leather-INCDTP; 2018. p. 33-7.",
"DOI": null,
"article": 99
},
{
"id": 3094,
"serial_number": 25,
"pmc": null,
"reference": "Camero M, Marinaro M, Lovero A, Elia G, Losurdo M, Buonavoglia C, et al. In vitro antiviral activity of Ficus carica latex against caprine herpesvirus-1. Nat Prod Res. 2014;28:2031-5.",
"DOI": null,
"article": 99
},
{
"id": 3095,
"serial_number": 26,
"pmc": null,
"reference": "Ghanbari A, Le Gresley A, Naughton D, Kuhnert N, Sirbu D, Ashrafi GH. Biological activities of Ficus carica latex for potential therapeutics in Human Papillomavirus (HPV) related cervical cancers. Sci Rep. 2019;9:1013.",
"DOI": null,
"article": 99
},
{
"id": 3096,
"serial_number": 27,
"pmc": null,
"reference": "Asl Najjari AH, Rajabi Z, Vasfi Marandi M, Dehghan G. The effect of the hexanic extracts of fig (Ficus carica) and olive (Olea europaea) fruit and nanoparticles of selenium on the immunogenicity of the inactivated avian influenza virus subtype H9N2. Vet Res Forum. 2015;6:227-31.",
"DOI": null,
"article": 99
},
{
"id": 3097,
"serial_number": 28,
"pmc": null,
"reference": "Ali MC, Munni YA, Das R, Sultana M, Akter N, Rahman M, et al. In silico chemical profiling and identification of neuromodulators from Curcuma amada targeting Acetylcholinesterase. bioRxiv. 2020.",
"DOI": null,
"article": 99
},
{
"id": 3098,
"serial_number": 29,
"pmc": null,
"reference": "Kim S, Chen J, Cheng T, Gindulyte A, He J, He S, et al. PubChem 2019 update: improved access to chemical data. Nucleic Acids Res. 2019;47:D1102-d9.",
"DOI": null,
"article": 99
},
{
"id": 3099,
"serial_number": 30,
"pmc": null,
"reference": "O’Boyle NM, Banck M, James CA, Morley C, Vandermeersch T, Hutchison GR. Open Babel: An open chemical toolbox. J Cheminformatics. 2011;3:33.",
"DOI": null,
"article": 99
},
{
"id": 3100,
"serial_number": 31,
"pmc": null,
"reference": "Dallakyan S, Olson AJ. Small-molecule library screening by docking with PyRx. Methods Mol Biol. 2015;1263:243-50.",
"DOI": null,
"article": 99
},
{
"id": 3101,
"serial_number": 32,
"pmc": null,
"reference": "Halgren TA. Merck molecular force field. I. Basis, form, scope, parameterization, and performance of MMFF94. J Comput Chem. 1996;17:490-519.",
"DOI": null,
"article": 99
},
{
"id": 3102,
"serial_number": 33,
"pmc": null,
"reference": "Burley SK, Berman HM, Bhikadiya C, Bi C, Chen L, Di Costanzo L, et al. RCSB Protein Data Bank: biological macromolecular structures enabling research and education in fundamental biology, biomedicine, biotechnology and energy. Nucleic Acids Res. 2019;47:D464-d74.",
"DOI": null,
"article": 99
},
{
"id": 3103,
"serial_number": 34,
"pmc": null,
"reference": "Guex N, Peitsch MC. SWISS‐MODEL and the Swiss‐Pdb Viewer: an environment for comparative protein modeling. Electrophoresis. 1997;18:2714-23.",
"DOI": null,
"article": 99
},
{
"id": 3104,
"serial_number": 35,
"pmc": null,
"reference": "Pettersen EF, Goddard TD, Huang CC, Couch GS, Greenblatt DM, Meng EC, et al. UCSF Chimera—a visualization system for exploratory research and analysis. J Comput Chem. 2004;25:1605-12.",
"DOI": null,
"article": 99
},
{
"id": 3105,
"serial_number": 36,
"pmc": null,
"reference": "Pagadala NS, Syed K, Tuszynski J. Software for molecular docking: a review. Biophys Rev. 2017;9:91-102.",
"DOI": null,
"article": 99
},
{
"id": 3106,
"serial_number": 37,
"pmc": null,
"reference": "Biovia DS. Discovery studio visualizer v4.5. Release; 2015.",
"DOI": null,
"article": 99
},
{
"id": 3107,
"serial_number": 38,
"pmc": null,
"reference": "Dickson CJ, Madej BD, Skjevik ÅA, Betz RM, Teigen K, Gould IR, et al. Lipid14: the amber lipid force field. Dickson CJ, Madej BD, Skjevik AA, et al. Lipid14: The Amber Lipid Force Field. J Chem Theory Comput. 2014;10(2):865-879. doi:10.10212014;10:865-79.",
"DOI": null,
"article": 99
},
{
"id": 3108,
"serial_number": 39,
"pmc": null,
"reference": "Krieger E, Vriend G, Spronk CJYo. YASARA–Yet Another Scientific Artificial Reality Application. 2013;993.",
"DOI": null,
"article": 99
},
{
"id": 3109,
"serial_number": 40,
"pmc": null,
"reference": "Dash R, Junaid M, Mitra S, Arifuzzaman M, Hosen SMZ. Structure-based identification of potent VEGFR-2 inhibitors from in vivo metabolites of a herbal ingredient. J Mol Model. 2019;25:98.",
"DOI": null,
"article": 99
},
{
"id": 3110,
"serial_number": 41,
"pmc": null,
"reference": "Mitra S, Dash R. Structural dynamics and quantum mechanical aspects of shikonin derivatives as CREBBP bromodomain inhibitors. J Mol Graph Model. 2018;83:42-52.",
"DOI": null,
"article": 99
},
{
"id": 3111,
"serial_number": 42,
"pmc": null,
"reference": "Dash R, Das R, Junaid M, Akash MF, Islam A, Hosen SZ. In silico-based vaccine design against Ebola virus glycoprotein. Adv appl bioinformatics chem. 2017;10:11-28.",
"DOI": null,
"article": 99
},
{
"id": 3112,
"serial_number": 43,
"pmc": null,
"reference": "Jorgensen WL, Chandrasekhar J, Madura JD, Impey RW, Klein MLJTJocp. Comparison of simple potential functions for simulating liquid water. J Chem Phy. 1983;79:926-35.",
"DOI": null,
"article": 99
},
{
"id": 3113,
"serial_number": 44,
"pmc": null,
"reference": "Krieger E, Nielsen JE, Spronk CA, Vriend GJJomg, modelling. Fast empirical pKa prediction by Ewald summation. J Mol Graph Model. 2006;25:481-6.",
"DOI": null,
"article": 99
},
{
"id": 3114,
"serial_number": 45,
"pmc": null,
"reference": "Krieger E, Vriend GJJocc. New ways to boost molecular dynamics simulations.J Comput Chem. 2015;36:996-1007.",
"DOI": null,
"article": 99
},
{
"id": 3115,
"serial_number": 46,
"pmc": null,
"reference": "Berendsen HJ, Postma Jv, van Gunsteren WF, DiNola A, Haak JRJTJocp. Molecular dynamics with coupling to an external bath. J Chem Phy. 1984;81:3684-90.",
"DOI": null,
"article": 99
},
{
"id": 3116,
"serial_number": 47,
"pmc": null,
"reference": "Humphrey W, Dalke A, Schulten KJJomg. VMD: visual molecular dynamics. J Mol Graph. 1996;14:33-8.",
"DOI": null,
"article": 99
},
{
"id": 3117,
"serial_number": 48,
"pmc": null,
"reference": "Dash R, Das R, Junaid M, Akash MFC, Islam A, Hosen SZJA, et al. In silico-based vaccine design against Ebola virus glycoprotein. Adv Appl Bioinform Chem. 2017;10:11.",
"DOI": null,
"article": 99
},
{
"id": 3118,
"serial_number": 49,
"pmc": null,
"reference": "Daina A, Michielin O, Zoete V. SwissADME: a free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci Rep. 2017;7:42717.",
"DOI": null,
"article": 99
},
{
"id": 3119,
"serial_number": 50,
"pmc": null,
"reference": "Lee S, Lee I, Kim H, Chang G, Chung J, No K. The PreADME Approach: Web-based program for rapid prediction of physico-chemical, drug absorption and drug-like properties. EuroQSAR. 2003.pp. 418-420.",
"DOI": null,
"article": 99
},
{
"id": 3120,
"serial_number": 51,
"pmc": null,
"reference": "Kontoyianni M. Docking and Virtual Screening in Drug Discovery. Methods Mol Biol. 2017;1647:255-66.",
"DOI": null,
"article": 99
},
{
"id": 3121,
"serial_number": 52,
"pmc": null,
"reference": "Neves BJ, Braga RC, Melo-Filho CC, Moreira-Filho JT, Muratov EN, Andrade CH. QSAR-Based Virtual Screening: Advances and Applications in Drug Discovery. FFront Pharmacol. 2018;9:1275.",
"DOI": null,
"article": 99
},
{
"id": 3122,
"serial_number": 53,
"pmc": null,
"reference": "Wang M, Cao R, Zhang L, Yang X, Liu J, Xu M, et al. Remdesivir and chloroquine effectively inhibit the recently emerged novel coronavirus (2019-nCoV) in vitro. Cell Res. 2020;30:269-71.",
"DOI": null,
"article": 99
},
{
"id": 3123,
"serial_number": 54,
"pmc": null,
"reference": "Wang Y, Zhang D, Du G, Du R, Zhao J, Jin Y, et al. Remdesivir in adults with severe COVID-19: a randomised, double-blind, placebo-controlled, multicentre trial. Lancet (London, England). 2020;395:1569-78.",
"DOI": null,
"article": 99
},
{
"id": 3124,
"serial_number": 55,
"pmc": null,
"reference": "Enmozhi SK, Raja K, Sebastine I, Joseph J. Andrographolide as a potential inhibitor of SARS-CoV-2 main protease: an in silico approach. J Biomol Struct Dyn. 2020:1-7.",
"DOI": null,
"article": 99
},
{
"id": 3125,
"serial_number": 56,
"pmc": null,
"reference": "Aref HL, Salah K, Chaumont JP, Fekih A, Aouni M, Said K. In vitro antimicrobial activity of four Ficus carica latex fractions against resistant human pathogens (antimicrobial activity of Ficus carica latex). Pak J Pharm Sci. 2010;23:53-8.",
"DOI": null,
"article": 99
},
{
"id": 3126,
"serial_number": 57,
"pmc": null,
"reference": "Dai W, Zhang B, Jiang XM, Su H, Li J, Zhao Y, et al. Structure-based design of antiviral drug candidates targeting the SARS-CoV-2 main protease. Science. 2020;368:1331-5.",
"DOI": null,
"article": 99
},
{
"id": 3127,
"serial_number": 58,
"pmc": null,
"reference": "Jin Z, Zhao Y, Sun Y, Zhang B, Wang H, Wu Y, et al. Structural basis for the inhibition of SARS-CoV-2 main protease by antineoplastic drug carmofur. Nat Struct Mol Biol. 2020;27:529-32.",
"DOI": null,
"article": 99
},
{
"id": 3128,
"serial_number": 59,
"pmc": null,
"reference": "Parvez MK, Alam P, Arbab AH, Al-Dosari MS, Alhowiriny TA, Alqasoumi SI. Analysis of antioxidative and antiviral biomarkers β-amyrin, β-sitosterol, lupeol, ursolic acid in Guiera senegalensis leaves extract by validated HPTLC methods. Saudi Pharm J. 2018;26:685-93.",
"DOI": null,
"article": 99
},
{
"id": 3129,
"serial_number": 60,
"pmc": null,
"reference": "Esposito F, Mandrone M, Del Vecchio C, Carli I, Distinto S, Corona A, et al. Multi-target activity of Hemidesmus indicus decoction against innovative HIV-1 drug targets and characterization of Lupeol mode of action. Pathog Dis. 2017;75.",
"DOI": null,
"article": 99
},
{
"id": 3130,
"serial_number": 61,
"pmc": null,
"reference": "Ahmad SF, Pandey A, Kour K, Bani S. Downregulation of pro-inflammatory cytokines by lupeol measured using cytometric bead array immunoassay. Phytother Res. 2010;24:9-13.",
"DOI": null,
"article": 99
},
{
"id": 3131,
"serial_number": 62,
"pmc": null,
"reference": "Tsai FS, Lin LW, Wu CR. Lupeol and Its Role in Chronic Diseases. Adv Exp Med Biol. 2016;929:145-75.",
"DOI": null,
"article": 99
},
{
"id": 3132,
"serial_number": 63,
"pmc": null,
"reference": "Krylova NV, Popov AM, Leonova GN, Artiukov AA, Maĭstrovskaia OS. [Comparative study of antiviral activity of luteolin and 7,3′-disulfate luteolin]. Antibiot Khimioter. 2011;56:7-10.",
"DOI": null,
"article": 99
},
{
"id": 3133,
"serial_number": 64,
"pmc": null,
"reference": "Fan W, Qian S, Qian P, Li X. Antiviral activity of luteolin against Japanese encephalitis virus. Virus Res. 2016;220:112-6.",
"DOI": null,
"article": 99
},
{
"id": 3134,
"serial_number": 65,
"pmc": null,
"reference": "Mehla R, Bivalkar-Mehla S, Chauhan A. A flavonoid, luteolin, cripples HIV-1 by abrogation of tat function. PloS one. 2011;6:e27915.",
"DOI": null,
"article": 99
},
{
"id": 3135,
"serial_number": 66,
"pmc": null,
"reference": "Wu S, Wang HQ, Guo TT, Li YH. Luteolin inhibits CVB3 replication through inhibiting inflammation. J Asian Nat Prod Res. 2019:1-12.",
"DOI": null,
"article": 99
},
{
"id": 3136,
"serial_number": 67,
"pmc": null,
"reference": "Dror RO, Dirks RM, Grossman JP, Xu H, Shaw DE. Biomolecular simulation: a computational microscope for molecular biology. Annu Rev Biophys. 2012;41:429-52.",
"DOI": null,
"article": 99
},
{
"id": 3137,
"serial_number": 68,
"pmc": null,
"reference": "Dash R, Choi HJ, Moon IS. Mechanistic insights into the deleterious roles of Nasu-Hakola disease associated TREM2 variants. Sci Rep. 2020;10:1-14.",
"DOI": null,
"article": 99
},
{
"id": 3138,
"serial_number": 69,
"pmc": null,
"reference": "Dash R, Ali MC, Dash N, Azad MAK, Hosen SMZ, Hannan MA, et al. Structural and Dynamic Characterizations Highlight the Deleterious Role of SULT1A1 R213H Polymorphism in Substrate Binding. Int J Mol Sci. 2019;20.",
"DOI": null,
"article": 99
},
{
"id": 3139,
"serial_number": 70,
"pmc": null,
"reference": "Talele TT, Khedkar SA, Rigby AC. Successful applications of computer aided drug discovery: moving drugs from concept to the clinic. Curr Top Med Chem. 2010;10:127-41.",
"DOI": null,
"article": 99
},
{
"id": 3140,
"serial_number": 71,
"pmc": null,
"reference": "Hirono S. [An introduction to the computer-aided structure-based drug design–applications of bioinformatics to drug discovery]. Jpn J Clin Pathol. 2002;50:45-51.",
"DOI": null,
"article": 99
},
{
"id": 3141,
"serial_number": 72,
"pmc": null,
"reference": "Ivanov AS, Veselovsky AV, Dubanov AV, Skvortsov VS. Bioinformatics platform development: from gene to lead compound. Methods Mol Biol. 2006;316:389-431.",
"DOI": null,
"article": 99
},
{
"id": 3142,
"serial_number": 73,
"pmc": null,
"reference": "Lipinski CA, Lombardo F, Dominy BW, Feeney PJ. Experimental and computational approaches to estimate solubility and permeability in drug discovery and development settings. Adv Drug Deliv. 1997;23:3-25.",
"DOI": null,
"article": 99
},
{
"id": 3143,
"serial_number": 74,
"pmc": null,
"reference": "Rahman A, Naheed NH, Raka SC, Qais N, Momen AZMR. Ligand-based virtual screening, consensus molecular docking, multi-target analysis and comprehensive ADMET profiling and MD stimulation to find out noteworthy tyrosine kinase inhibitor with better efficacy and accuracy. Orient Pharm Exp Med. 2019. doi.org/10.1007/s13596-019-00406-9.",
"DOI": null,
"article": 99
}
]
},
{
"id": 91,
"slug": "178-1594574470-prediction-of-b-cell-and-t-cell-epitopes-in-the-spike-glycoprotein-of-sars-cov-2-in-bangladesh-an-in-silico-approach",
"featured": false,
"slider": false,
"issue": "Special Issue",
"type": "original_article",
"manuscript_id": "178-1594574470",
"recieved": "2020-07-12",
"revised": null,
"accepted": "2020-08-21",
"published": "2020-09-02",
"pdf_file": "https://jabet.bsmiab.org/media/pdf_file/2023/07/178-1594574470.pdf",
"title": "Prediction of B-cell and T-cell epitopes in the spike glycoprotein of SARS-CoV-2 in Bangladesh: An in-silico approach",
"abstract": "<p>To combat highly infectious Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2), scientists and researchers are toiling hard globally to develop effective drugs and vaccines. By exploring the structural proteins of SARS-CoV-2 can be a feasible way to find an effective vaccine. In this study by using in-silico tools, we recommended B-cell and T-cell epitopes of spike protein from a Bangladeshi isolate which can be considered for incorporation into a vaccine against the SARS-CoV-2. Homology modelling, energy minimization process, and finally Ramachandran model was used for the prediction of a more stable conformation of the spike protein. The most important peptides were screened through the VaxiJen server followed by the IEDB server and CTLPred Score predicted and analysed the desired epitopes. In the final analysis, the peptide EVRQIAPGQTGKIADY (starting from 91) showed the highest antigenicity score (1.3837) as a B-cell epitope although GSTPCNGVEGFNCYFP, starting at 161, showed highest score (0.91) in an initial analysis. On the contrary, as a T-cell epitope, 71 KLNDLCFTNV- 80 was found with the highest antigenicity score (2.6927) which was also found as an epitope in further analysis. A combination of B-cell and T-cell epitopes may evoke a humoral and cell-mediated immune response which will possibly lead to an effective vaccine. Further, the various computational analyses will provide valuable information that will pave the way for modelling a novel vaccine against SARS-CoV-2.</p>",
"journal_reference": "J Adv Biotechnol Exp Ther. 2020; 3(4): 49-56.",
"academic_editor": "Md Abdul Hannan, PhD; Dongguk University, South Korea",
"cite_info": "Islam MA, Shihab MMR, et al. Prediction of B-cell and T-cell epitopes in the spike glycoprotein of SARS-CoV-2 in Bangladesh: An in-silico approach. J Adv Biotechnol Exp Ther. 2020; 3(4): 49-56.",
"keywords": [
"SARS-CoV-2",
"COVID-19",
"Bangladesh",
"B-cell epitope",
"T-cell epitope"
],
"DOI": "10.5455/jabet.2020.d156",
"sections": [
{
"section_number": 1,
"section_title": "INTRODUCTION",
"body": "<p>Coronavirus disease 2019 (COVID-19) pandemic is the consequence of the respiratory tract infection caused by novel Coronavirus (2019‐nCoV) which is later known as Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2). It has emerged on the eve of 2020 from the Chinese city of Wuhan (Hubei province) [<a href=\"#r-1\">1</a>]. As of June 15, 2020, the pandemic has globally claimed more than 436,900 lives and infected over 8.03 million individuals [<a href=\"#r-2\">2</a>]. Therefore, the development of a vaccine is no longer merely a discussion or part of a debate about whether a vaccine is ultimately needed to suppress the viral spread and to control and prevent SARS-CoV-2. As of June 29, 2020, seventeen candidate vaccines were in clinical evaluation across the globe whereas other 132 candidates in preclinical stages [<a href=\"#r-3\">3</a>].<br />\r\nSARS-CoV-2, which is a positive‐sense single‐stranded RNA virus, has a genome size of approximately 30 kilobases which encode for various structural and non‐structural proteins. The structural proteins of SARS-CoV-2 are the spike (S) protein, the envelope (E) protein, the membrane (M) protein, and the nucleocapsid (N) protein and the non-structural proteins contain open reading frame 1ab (ORF1ab), ORF3a, ORF6, ORF7a, ORF8, and ORF10 [<a href=\"#r-4\">4, 5</a>]. The S protein has two major domains: S1 and S2 [<a href=\"#r-6\">6</a>]. S1 subunit has the receptor-binding domain (RBD) and binds to angiotensin-converting enzyme 2 (ACE2) while S2 mediates the fusion of viral and host cell membranes [<a href=\"#r-7\">7, 8</a>].<br />\r\nThe first laboratory-confirmed case in Bangladesh was identified on March 8, 2020 [<a href=\"#r-9\">9</a>]. So far, 90,619 people have been tested positive and 1,209 have died in the country (as of June 15, 2020) [<a href=\"#r-10\">10</a>]. Genomic data of SARS-CoV-2 in Bangladesh is limited, however, efforts from scientists here are underway to make genome sequences available gradually [<a href=\"#r-11\">11</a>]. On May 11, 2020, the first whole-genome sequence (EPI_ISL_437912) from Bangladesh was published. A few days later, the study sequence (EPI_ISL_447590) was made publicly available by the Genome Centre, Jashore University of Science and Technology, Jashore, Bangladesh.<br />\r\nTo develop an effective vaccine, the identification of B-cell and T-cell epitopes for SARS-CoV-2 proteins are critical especially for S protein. Both humoral immunity and cellular immunity, provided by B-cell antibodies and T-cell respectively, are essential for effective vaccines [<a href=\"#r-12\">12, 13</a>]. By analyzing an S protein of SARS‐CoV‐2 from Bangladesh, we identified and reported the B-cell and T-cell epitopes. Therefore, the epitopes presented in the study may evoke an effective immune response against SARS‐CoV‐2 for which further experimental analysis will be required. We tend to target the epitopes within the S protein as the protein gave a dominant and long‐lasting immune response against SARS‐CoV that was previously reported [<a href=\"#r-4\">4</a>]. However, our analyses will probably slender down the rummage around for targets for designing a novel vaccine candidate against SARS-CoV-2.</p>"
},
{
"section_number": 2,
"section_title": "MATERIALS AND METHODS",
"body": "<p><strong>Nucleotide sequence retrieval and structure prediction</strong><br />\r\nThe nucleotide sequence (EPI_ISL_447590) of a Bangladeshi SARS‐CoV‐2 spike (S) glycoprotein was downloaded from the GISAID EpiCoV<sup>TM</sup> database. The nucleotide sequence was then translated into an amino acid sequence for further analysis. The protein sequence was then applied to form its 3-D model by using SWISS-MODEL [<a href=\"#r-14\">14</a>]. The SMTL ID: 6vyb.1 (<a href=\"https://swissmodel.expasy.org/templates/6vyb.1\">https://swissmodel.expasy.org/templates/6vyb.1</a>), chain A, was used as a template that shared 100% sequence identity with the study sequence. The 6vyb.1, chain A, is a SARS-CoV-2 spike ectodomain structure (open state) solved experimentally using Electron Microscopy deposited at early this year in the protein data bank [<a href=\"#r-15\">15</a>]. A PDB file was found after the modeling which was then used for further prediction. Moreover, we minimized energy after homology modeling to find out a stable conformation of proteins using FoldX (<a href=\"http://foldxsuite.crg.eu/\">http://foldxsuite.crg.eu/</a>). Ramachandran model was also used for finding stable conformation. Ramachandran plot that we used here to determine torsional angles which are permitted, so we could understand the insight into the structure of the peptides, ultimately stable conformation of the peptides. In the Ramachandran plot, the outlier with percentage was measured by SWISS-PDB. From the outliers, the conformations of the phi (Φ) and psi (Ψ) angles were predicted in the protein. Moreover, empirical distribution was observed in this single structure. Ultimately protein stability was confirmed by this value.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Prediction of B-cell epitope</strong><br />\r\nThe linear and discontinuous B-cell epitopes based on protein’s 3-D structure were predicted using Ellipro [<a href=\"#r-16\">16</a>]. The prediction parameters were a minimum score and a maximum distance of 0.5 and 6, respectively. The number of residues of each chain was predicted by Ellipro. For identifying B-cell epitopes in an antigen sequence, ABCpred was used to identify the peptide sequence [<a href=\"#r-17\">17</a>]. The protein sequence was submitted where the threshold was 0.51 with a length of 16 to be used for prediction. The antigenicity index of the predicted epitopes was examined and for predicting and designing Interferon-gamma (IFN-γ) inducing peptides, a combination of VaxiJen v2.0 with a threshold of 0.4 and IFN-γ response were used [<a href=\"#r-18\">18, 19</a>].</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Prediction of T-cell epitope</strong><br />\r\nThe T-cell epitope prediction was performed by using the IEDB tool where all MHC alleles were taken with a threshold of 0.7 for the study [<a href=\"#r-20\">20-22</a>]. The output peptides were again examined by using a combination of Vixen, IFN-γ response, and CTLPred which scores were based on using both support vector machine (SVM) and artificial neural network (ANN) score [<a href=\"#r-23\">23</a>]. In CTLPred, ANN and SVM cut-off scores were used as of 0.51 and 0.36, respectively.</p>"
},
{
"section_number": 3,
"section_title": "RESULTS",
"body": "<p><strong>Nucleotide sequence retrieval and structure prediction</strong><br />\r\nThe retrieved nucleotide sequence from the GISAID EpiCoV<sup>TM</sup> database was found to have a length of 694 base pairs. The converted protein sequence was then used for modeling (<a href=\"#figure1\">Figure 1</a>) which showed a 100% sequence identity with template sequence. After conducting the energy minimization process, we got energy minimized state of that protein (<a href=\"#Table-1\">Table 1</a>) and a new 3-D figure of the S protein (<a href=\"#figure2\">Figure 2</a>).<br />\r\nFrom the Ramachandran model (<a href=\"#figure3\">Figure 3</a>), the stable conformation of the PDB file was confirmed with Ramachandran Favoured, and Ramachandran Outliers of 92.58%, and 1.75%, respectively.</p>\r\n\r\n<div id=\"figure1\">\r\n<figure class=\"image\"><img alt=\"\" height=\"303\" src=\"/media/article_images/2024/03/07/178-1594574470-Figure1.jpg\" width=\"335\" />\r\n<figcaption><strong>Figure 1</strong>. Homology modeling of spike protein generated by SWISS-MODEL.</figcaption>\r\n</figure>\r\n</div>\r\n\r\n<div id=\"figure2\">\r\n<figure class=\"image\"><img alt=\"\" height=\"278\" src=\"/media/article_images/2024/03/07/178-1594574470-Figure2.jpg\" width=\"334\" />\r\n<figcaption><strong>Figure 2.</strong> 3-D model of the spike protein after energy optimization generated by using FoldX.</figcaption>\r\n</figure>\r\n</div>\r\n\r\n<div id=\"figure3\">\r\n<figure class=\"image\"><img alt=\"\" height=\"308\" src=\"/media/article_images/2024/03/07/178-1594574470-Figure3.jpg\" width=\"336\" />\r\n<figcaption><strong>Figure 3. </strong>Ramachandran plot using SWISS-MODEL.</figcaption>\r\n</figure>\r\n</div>\r\n\r\n<div id=\"Table-1\">\r\n<p><a href=\"https://jabet.bsmiab.org/table/178-1594574470-table1/\">Table-1</a><strong>Table 1.</strong> Energy minimization of spike protein sequence to find out a stable conformation of protein.</p>\r\n</div>\r\n\r\n<p> </p>\r\n\r\n<p><strong>B-cell epitope prediction</strong><br />\r\nThe PDB file was used to identify linear and discontinuous epitopes by using Ellipro. This revealed the presence of B-cell epitopes according to their chain ID. It showed 3 chain IDs with their number of residues, 231 equally (<a href=\"#figure4\">Figure 4</a>).<br />\r\nThe linear peptides sequence with a threshold of 0.51 provided 23 sequences at a specific length of 16. The antigenicity of peptides was confirmed by the score (<a href=\"#Table-2\">Table 2</a>).<br />\r\nFor more confirmation, combined processes were done to identify antigen in the spike glycoprotein (<a href=\"#Table-3\">Table 3</a>). IFN-γ responses can be either positive or negative depending on the cytokines. The probability of being an antigen of specific peptides was confirmed by VaxiJen with a threshold of 0.4.</p>\r\n\r\n<div id=\"figure4\">\r\n<figure class=\"image\"><img alt=\"\" height=\"190\" src=\"/media/article_images/2024/03/07/178-1594574470-Figure4.jpg\" width=\"336\" />\r\n<figcaption><strong>Figure 4. </strong>Predicted linear B-cell epitopes and their locations to the surface membrane (yellow) by Ellipro.</figcaption>\r\n</figure>\r\n</div>\r\n\r\n<div id=\"Table-2\">\r\n<p><a href=\"https://jabet.bsmiab.org/table/178-1594574470-table2/\">Table-2</a><strong>Table 2. </strong>Predicted antigenic B-cell linear epitopes found in the spike glycoprotein with their positions and antigenicity scores.</p>\r\n</div>\r\n\r\n<div id=\"Table-3\">\r\n<p><a href=\"https://jabet.bsmiab.org/table/178-1594574470-table3/\">Table-3</a><strong>Table 3. </strong>Predicted antigenic B-cell linear epitopes found in the spike glycoprotein with their antigenic scores, VaxiJen scores, and IFN-γ responses.</p>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Prediction of T-cell epitope</strong><br />\r\nIn spike glycoprotein, a total of 27 MHC-I peptides were predicted at different amino acid positions. However, 12 peptide sequences were found to be antigenic in VaxiJen. IFN-γ responses can be either positive or negative. MHC restriction refers that a particular MHC molecule bounded with the peptides that can be interacted with T cell epitopes. CTLPred score confirms the epitopes probability by combining both support vector machine (SVM) and artificial neural network (ANN) score (<a href=\"#Table-4\">Table 4</a>).</p>\r\n\r\n<div id=\"Table-4\">\r\n<p><a href=\"https://jabet.bsmiab.org/table/178-1594574470-table4/\">Table-4</a><strong>Table 4.</strong> MHC-I associated antigenic peptide predicted on the spike glycoprotein.</p>\r\n</div>"
},
{
"section_number": 4,
"section_title": "DISCUSSION",
"body": "<p>To control the current pandemic, the development of new drugs and vaccines is an urgent issue. Especially, effective vaccination or immunotherapy could play a remarkable role in suppressing the viral spread and eventually eliminating it from humans. In the case of vaccine development, a challenge is the absence of sufficient data concerning specific immune responses against SARS‐CoV‐2.<br />\r\nThis study has recommended some epitopes that may have the ability to evoke sufficient response in the human body and to be incorporated into a novel vaccine. The 3-D structure of spike (S) protein derived from a Bangladeshi SARS-CoV-2 strain was firstly determined, minimized energy, and confirmed by the Ramachandran model followed by the probable antigenic B-cell, and T-cell epitopes prediction using the amino acid sequence of the same protein.<br />\r\nRecently, a report unveiled that T-cell epitopes are more conducive as a result of more long-lasting immune response mediated by CD8+ T-cell and due to the antigenic drift, by which an antibody is not able to respond against an antibody [24]. In this study, we predicted 12 peptides as T-cell epitopes in a combined strategy (VaxiJen score and IFN-γ responses). All of the peptides were antigenic whereas 71- KLNDLCFTNV- 80 showed the highest antigenicity score (2.6927) though the IFN-γ response was negative which mainly depends on cytokines and can be either positive or negative. In another analysis (CTLPred score), the peptide was also found as an epitope with support vector machine (SVM) and artificial neural network (ANN) score was 0.03 and 0.67438, respectively. Besides these, all peptides were able to interact with the MHC class I alleles.<br />\r\nIn the case of SARS‐CoV, antibodies were generated in mice against spike protein that gave protection from the infection [<a href=\"#r-25\">25-27</a>]. Besides, the B-cell epitope provides a strong immune response without causing any adverse effects [<a href=\"#r-28\">28</a>]. Therefore, we also predicted 23 linear B-cell epitopes of 16-mer which could be effective. Among these peptides, the peptide GSTPCNGVEGFNCYFP with start position 161 showed the highest antigenicity score (0.91). But further analysis (VaxiJen score and IFN-γ responses) revealed only 13 peptides and among them, the peptide EVRQIAPGQTGKIADY, started from 91, showed the highest antigenicity score (1.3837) with negative (1) IFN-γ responses.<br />\r\nThe potential candidate epitopes, reported in this study, may contribute to the development of a novel vaccine against SARS-CoV-2. A large number of Bangladeshi individuals can be covered by these peptides. The limitation of the study is that all the analyses here were based on a single sequence of SARS‐CoV‐2 derived from Bangladesh. More mutations will be observed as the virus is evolving continuously which may affect our present analysis although most of the mutations are synonymous. However, further experimental analysis will also be required to prove the immunogenicity of the recommended peptides. Overall, the study provides insight that will eventually contribute to the development of a vaccine against SARS‐CoV‐2.</p>"
},
{
"section_number": 5,
"section_title": "ACKNOWLEDGEMENT",
"body": "<p>This research received no external funding. We are grateful to the authors and the laboratory from where the study sequence was originated and submitted followed by sharing through the GISAID EpiCoV<sup>TM </sup>database. Accession ID of the study sequence: EPI_ISL_447590 Originating and submitting laboratory: Genome Centre, Jashore University of Science and Technology, Jashore-7408, Bangladesh. Authors: A. S. M. Rubayet Ul Alam, M. Rafiul Islam, M. Shaminur Rahman, Md. Tanvir Islam, Md. Shazid Hasan, Pravas Chandra Roy, Habiba Ibnat, MD. Ali Ahasan Setu, Tanay Chakrovarty, Sourav Dutta Dip, Ruhul Amin, Md Nur Kabidul Azam, Ovinu Kibria Islam, Hassan M. Al-Emran, Shireen Nigar, Selina Akter, Md. Nazmul Hasan, Iqbal Kabir Jahid, M. Anwar Hossain.</p>"
},
{
"section_number": 6,
"section_title": "CONFLICT OF INTEREST",
"body": "<p>The authors declare that there is no conflict of interest.</p>"
},
{
"section_number": 7,
"section_title": "AUTHOR CONTRIBUTIONS",
"body": "<p>MAI conceived the idea and supervised the project. MH performed the database search. MMRS contributed to perform the experiments. MH analyzed data and wrote the manuscript. MH and MMRS illustrated the figures and prepared the tables. MAI critically revised the manuscript. All the authors proofread and approved the final manuscript.</p>"
}
],
"figures": [
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/03/07/178-1594574470-Figure1.jpg",
"caption": "Figure 1. Homology modeling of spike protein generated by SWISS-MODEL.",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/03/07/178-1594574470-Figure2.jpg",
"caption": "Figure 2. 3-D model of the spike protein after energy optimization generated by using FoldX.",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/03/07/178-1594574470-Figure3.jpg",
"caption": "Figure 3. Ramachandran plot using SWISS-MODEL.",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2024/03/07/178-1594574470-Figure4.jpg",
"caption": "Figure 4. Predicted linear B-cell epitopes and their locations to the surface membrane (yellow) by Ellipro.",
"featured": false
}
],
"authors": [
{
"id": 341,
"affiliation": [
{
"affiliation": "Department of Microbiology, Faculty of Life and Earth Science, Jagannath University, Dhaka 1100, Bangladesh"
}
],
"first_name": "Mehedi",
"family_name": "Hasan",
"email": null,
"author_order": 1,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 91
},
{
"id": 342,
"affiliation": [
{
"affiliation": "Department of Microbiology, Faculty of Life and Earth Science, Jagannath University, Dhaka 1100, Bangladesh"
}
],
"first_name": "Md. Maksudur Rahman",
"family_name": "Shihab",
"email": null,
"author_order": 2,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 91
},
{
"id": 343,
"affiliation": [
{
"affiliation": "Department of Microbiology, Faculty of Life and Earth Science, Jagannath University, Dhaka 1100, Bangladesh"
}
],
"first_name": "Mohammad Ariful",
"family_name": "Islam",
"email": "ariful@mib.jnu.ac.bd",
"author_order": 3,
"ORCID": null,
"corresponding": true,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "Mohammad Ariful Islam, PhD; Department of Microbiology, Faculty of Life and Earth Science, Jagannath University, Dhaka 1100, Bangladesh, Email: ariful@mib.jnu.ac.bd",
"article": 91
}
],
"views": 767,
"downloads": 145,
"references": [
{
"id": 2716,
"serial_number": 1,
"pmc": null,
"reference": "Wang C, Horby PW, Hayden FG, Gao GF. A novel coronavirus outbreak of global health concern. The Lancet. 2020;395(10223):470-3.",
"DOI": null,
"article": 91
},
{
"id": 2717,
"serial_number": 2,
"pmc": null,
"reference": "Dong E, Du H, Gardner L. An interactive web-based dashboard to track COVID-19 in real time. The Lancet infectious diseases. 2020;20(5):533-4.",
"DOI": null,
"article": 91
},
{
"id": 2718,
"serial_number": 3,
"pmc": null,
"reference": "World Health Organization (WHO). DRAFT landscape of COVID-19 candidate vaccines – 29 June 2020. Retrieved June 30, 2020, from https://www.who.int/publications/m/item/draft-landscape-of-covid-19-candidate-vaccines.",
"DOI": null,
"article": 91
},
{
"id": 2719,
"serial_number": 4,
"pmc": null,
"reference": "Ahmed SF, Quadeer AA, McKay MR. Preliminary identification of potential vaccine targets for the COVID-19 coronavirus (SARS-CoV-2) based on SARS-CoV immunological studies. Viruses. 2020;12(3):254.",
"DOI": null,
"article": 91
},
{
"id": 2720,
"serial_number": 5,
"pmc": null,
"reference": "Kiyotani K, Toyoshima Y, Nemoto K, Nakamura Y. Bioinformatic prediction of potential T cell epitopes for SARS-Cov-2. Journal of Human Genetics. 2020;65(7):569-75.",
"DOI": null,
"article": 91
},
{
"id": 2721,
"serial_number": 6,
"pmc": null,
"reference": "Ortega JT, Serrano ML, Pujol FH, Rangel HR. Role of changes in SARS-CoV-2 spike protein in the interaction with the human ACE2 receptor: An in silico analysis. EXCLI journal. 2020; 19:410.",
"DOI": null,
"article": 91
},
{
"id": 2722,
"serial_number": 7,
"pmc": null,
"reference": "Hoffmann M, Kleine-Weber H, Schroeder S, Krüger N. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is 278 Blocked by a Clinically Proven Protease Inhibitor. Cell. 2020. doi: 10.1016/j.cell.2020.02.052.",
"DOI": null,
"article": 91
},
{
"id": 2723,
"serial_number": 8,
"pmc": null,
"reference": "Verdecchia P, Cavallini C, Spanevello A, Angeli F. The pivotal link between ACE2 deficiency and SARS-CoV-2 infection. European Journal of Internal Medicine. 2020. doi: 10.1016/j.ejim.2020.04.037.",
"DOI": null,
"article": 91
},
{
"id": 2724,
"serial_number": 9,
"pmc": null,
"reference": "Hossain I, Khan MH, Rahman MS, Mullick AR, Aktaruzzaman M. The epidemiological characteristics of an outbreak of 2019 novel coronavirus diseases (COVID-19) in Bangladesh: A descriptive study. Journal of Medical Science and Clinical Research. 2020;8(04).",
"DOI": null,
"article": 91
},
{
"id": 2725,
"serial_number": 10,
"pmc": null,
"reference": "WHO Bangladesh COVID-19 Situation Report #16, 15 June 2020. Retrieved June 18, 2020, from https://www.who.int/bangladesh/emergencies/coronavirus-disease-(covid-19)-update/coronavirus-disease-(covid-2019)-bangladesh-situation-reports.",
"DOI": null,
"article": 91
},
{
"id": 2726,
"serial_number": 11,
"pmc": null,
"reference": "Hasan S, Khan S, Ahsan GU, Hossain MM. Genome Analysis of SARS-CoV-2 Isolate from Bangladesh. BioRxiv. 2020. doi:10.1101/2020.05.13.094441.",
"DOI": null,
"article": 91
},
{
"id": 2727,
"serial_number": 12,
"pmc": null,
"reference": "Olsson S-E, Villa LL, Costa RL, Petta CA, Andrade RP, Malm C, et al. Induction of immune memory following administration of a prophylactic quadrivalent human papillomavirus (HPV) types 6/11/16/18 L1 virus-like particle (VLP) vaccine. Vaccine. 2007;25(26):4931-9.",
"DOI": null,
"article": 91
},
{
"id": 2728,
"serial_number": 13,
"pmc": null,
"reference": "Rappuoli R, Black S, Bloom DE. Vaccines and global health: In search of a sustainable model for vaccine development and delivery. Science Translational Medicine. 2019;11(497): eaaw2888.",
"DOI": null,
"article": 91
},
{
"id": 2729,
"serial_number": 14,
"pmc": null,
"reference": "Waterhouse A, Bertoni M, Bienert S, Studer G, Tauriello G, Gumienny R, et al. SWISS-MODEL: homology modelling of protein structures and complexes. Nucleic acids research. 2018;46(W1): W296-W303.",
"DOI": null,
"article": 91
},
{
"id": 2730,
"serial_number": 15,
"pmc": null,
"reference": "Walls AC, Park Y-J, Tortorici MA, Wall A, McGuire AT, Veesler D. Structure, function, and antigenicity of the SARS-CoV-2 spike glycoprotein. Cell. 2020. doi: 10.1016/j.cell.2020.02.058.",
"DOI": null,
"article": 91
},
{
"id": 2731,
"serial_number": 16,
"pmc": null,
"reference": "Ponomarenko J, Bui H-H, Li W, Fusseder N, Bourne PE, Sette A, et al. ElliPro: a new structure-based tool for the prediction of antibody epitopes. BMC bioinformatics. 2008;9(1):514.",
"DOI": null,
"article": 91
},
{
"id": 2732,
"serial_number": 17,
"pmc": null,
"reference": "Saha S, Raghava GPS. Prediction of continuous B‐cell epitopes in an antigen using recurrent neural network. Proteins: Structure, Function, and Bioinformatics. 2006;65(1):40-8.",
"DOI": null,
"article": 91
},
{
"id": 2733,
"serial_number": 18,
"pmc": null,
"reference": "Dhanda SK, Vir P, Raghava GP. Designing of interferon-gamma inducing MHC class-II binders. Biology direct. 2013;8(1):30.",
"DOI": null,
"article": 91
},
{
"id": 2734,
"serial_number": 19,
"pmc": null,
"reference": "Kearse M, Moir R, Wilson A, Stones-Havas S, Cheung M, Sturrock S, et al. Geneious Basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics. 2012;28(12):1647-9.",
"DOI": null,
"article": 91
},
{
"id": 2735,
"serial_number": 20,
"pmc": null,
"reference": "Andreatta M, Nielsen M. Gapped sequence alignment using artificial neural networks: application to the MHC class I system. Bioinformatics. 2016;32(4):511-7.",
"DOI": null,
"article": 91
},
{
"id": 2736,
"serial_number": 21,
"pmc": null,
"reference": "Lundegaard C, Lund O, Nielsen M. Accurate approximation method for prediction of class I MHC affinities for peptides of length 8, 10 and 11 using prediction tools trained on 9mers. Bioinformatics. 2008;24(11):1397-8.",
"DOI": null,
"article": 91
},
{
"id": 2737,
"serial_number": 22,
"pmc": null,
"reference": "Lundegaard C, Nielsen M, Lund O. The validity of predicted T-cell epitopes. Trends in biotechnology. 2006;24(12):537-8.",
"DOI": null,
"article": 91
},
{
"id": 2738,
"serial_number": 23,
"pmc": null,
"reference": "Bhasin M, Raghava G. Prediction of CTL epitopes using QM, SVM and ANN techniques. Vaccine. 2004;22(23-24):3195-204.",
"DOI": null,
"article": 91
},
{
"id": 2739,
"serial_number": 24,
"pmc": null,
"reference": "Chiou S-S, Fan Y-C, Crill WD, Chang R-Y, Chang G-JJ. Mutation analysis of the cross-reactive epitopes of Japanese encephalitis virus envelope glycoprotein. Journal of general virology. 2012;93(6):1185-92.",
"DOI": null,
"article": 91
},
{
"id": 2740,
"serial_number": 25,
"pmc": null,
"reference": "Deming D, Sheahan T, Heise M, Yount B, Davis N, Sims A, et al. Vaccine efficacy in senescent mice challenged with recombinant SARS-CoV bearing epidemic and zoonotic spike variants. PLoS medicine. 2006;3(12).",
"DOI": null,
"article": 91
},
{
"id": 2741,
"serial_number": 26,
"pmc": null,
"reference": "Graham RL, Becker MM, Eckerle LD, Bolles M, Denison MR, Baric RS. A live, impaired-fidelity coronavirus vaccine protects in an aged, immunocompromised mouse model of lethal disease. Nature medicine. 2012;18(12):1820.",
"DOI": null,
"article": 91
},
{
"id": 2742,
"serial_number": 27,
"pmc": null,
"reference": "Yang Z-y, Kong W-p, Huang Y, Roberts A, Murphy BR, Subbarao K, et al. A DNA vaccine induces SARS coronavirus neutralization and protective immunity in mice. Nature. 2004;428(6982):561-4.",
"DOI": null,
"article": 91
},
{
"id": 2743,
"serial_number": 28,
"pmc": null,
"reference": "Rakib A, Sami SA, Islam MA, Ahmed S, Faiz FB, Khanam BH, et al. Epitope-Based Peptide Vaccine Against Severe Acute Respiratory Syndrome-Coronavirus-2 Nucleocapsid Protein: An in silico Approach. BioRxiv. 2020. doi: 10.1101/2020.05.16.100206.",
"DOI": null,
"article": 91
}
]
}
]
}
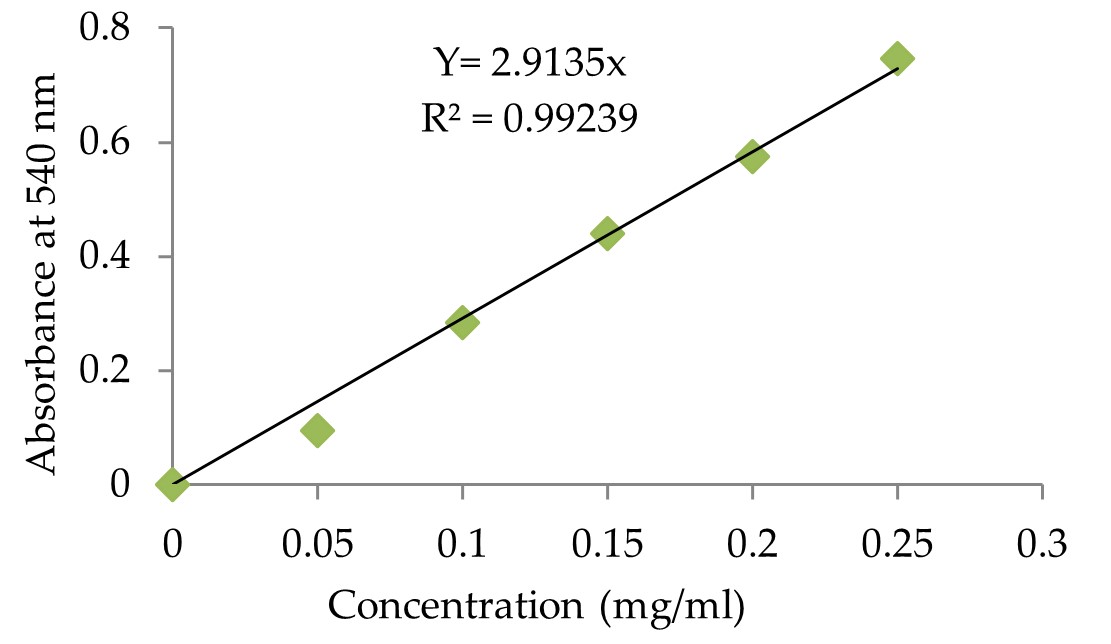{kind=link}
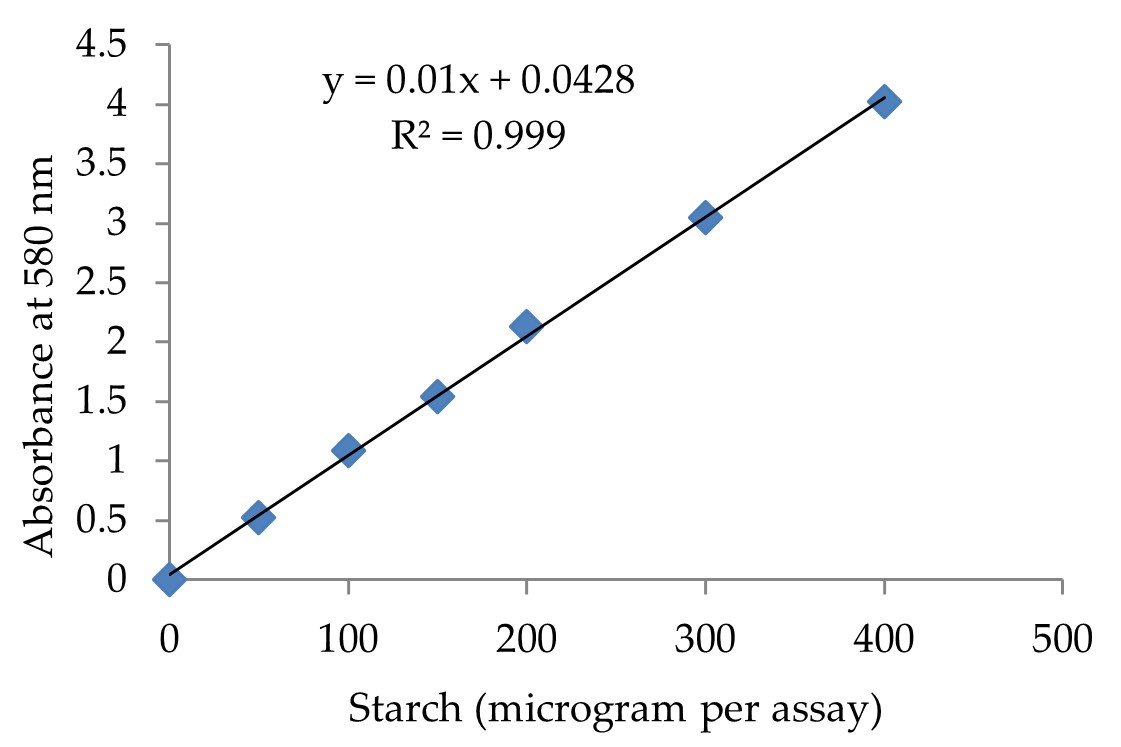{kind=link}
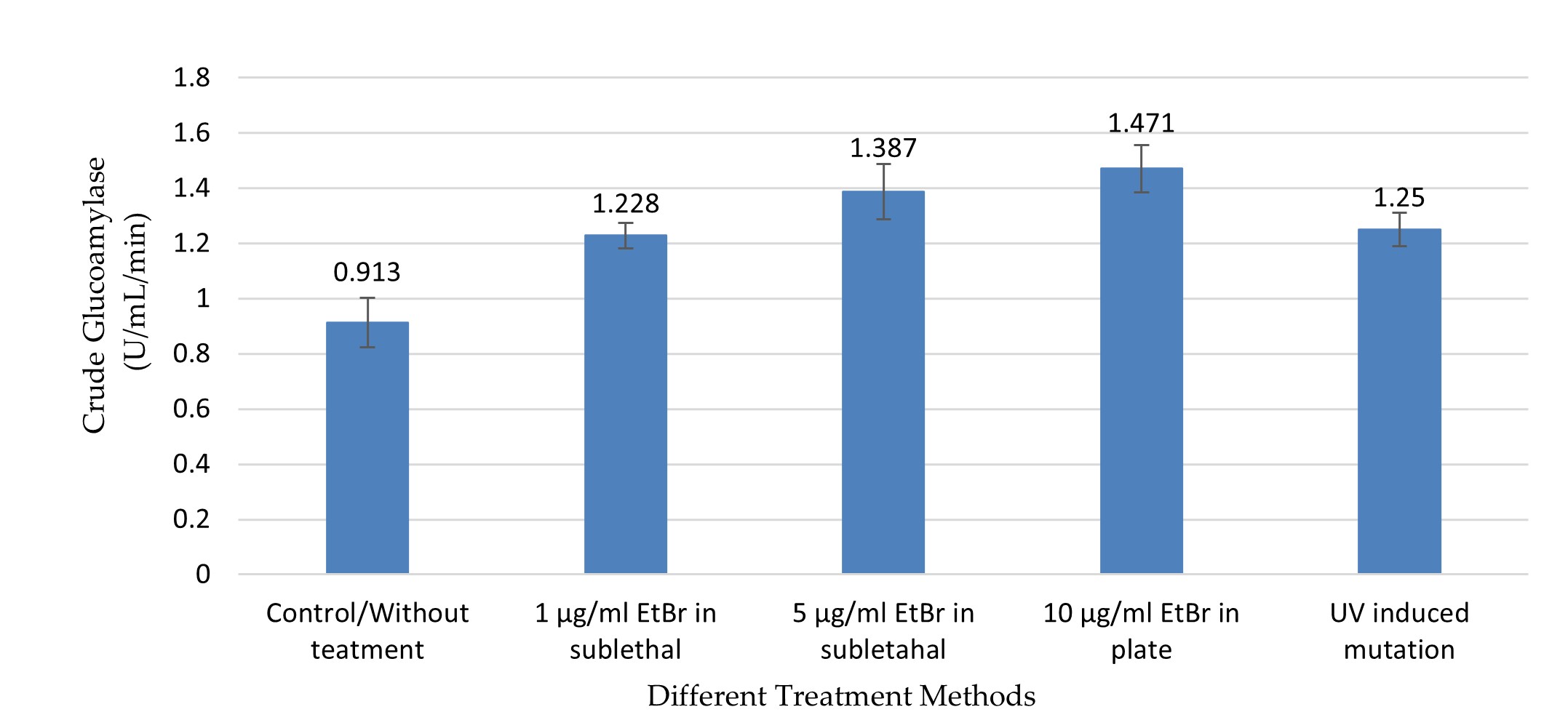{kind=link}
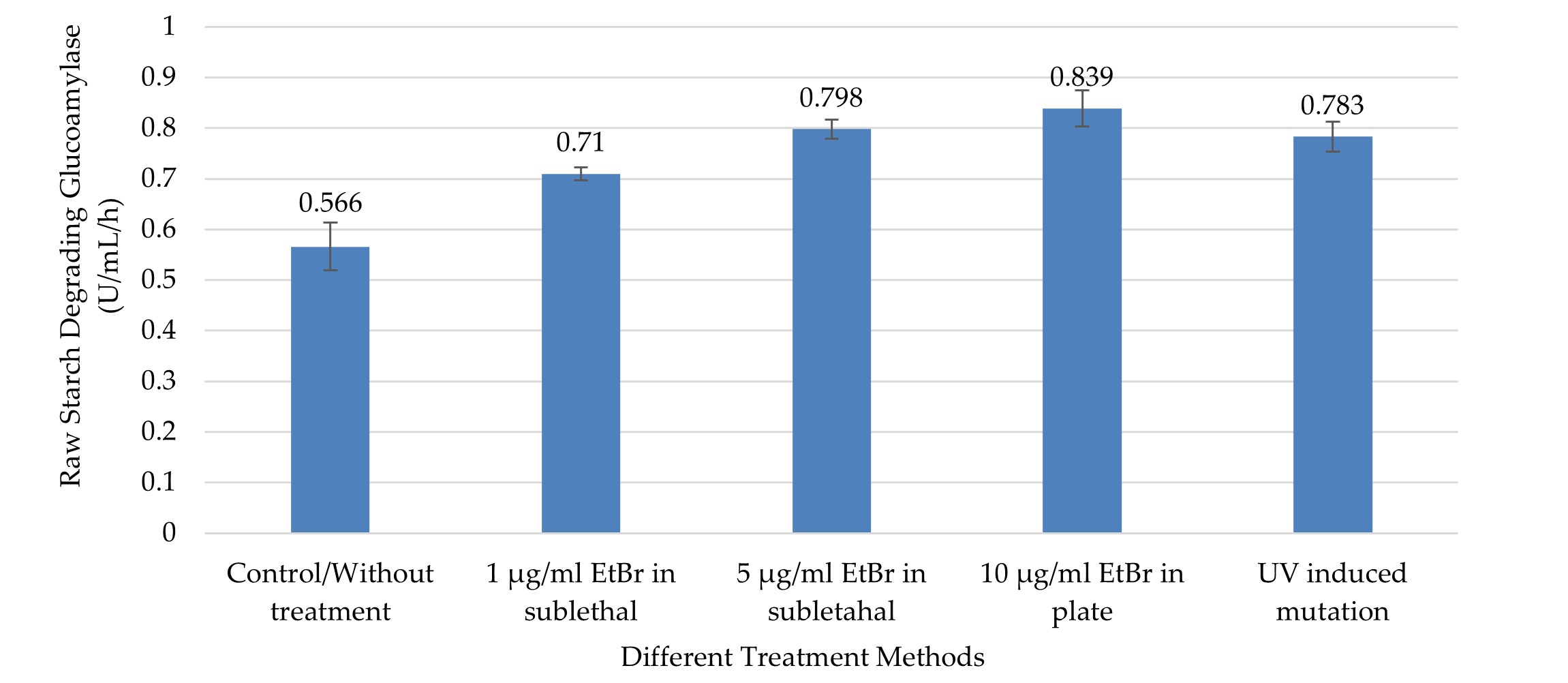{kind=link}
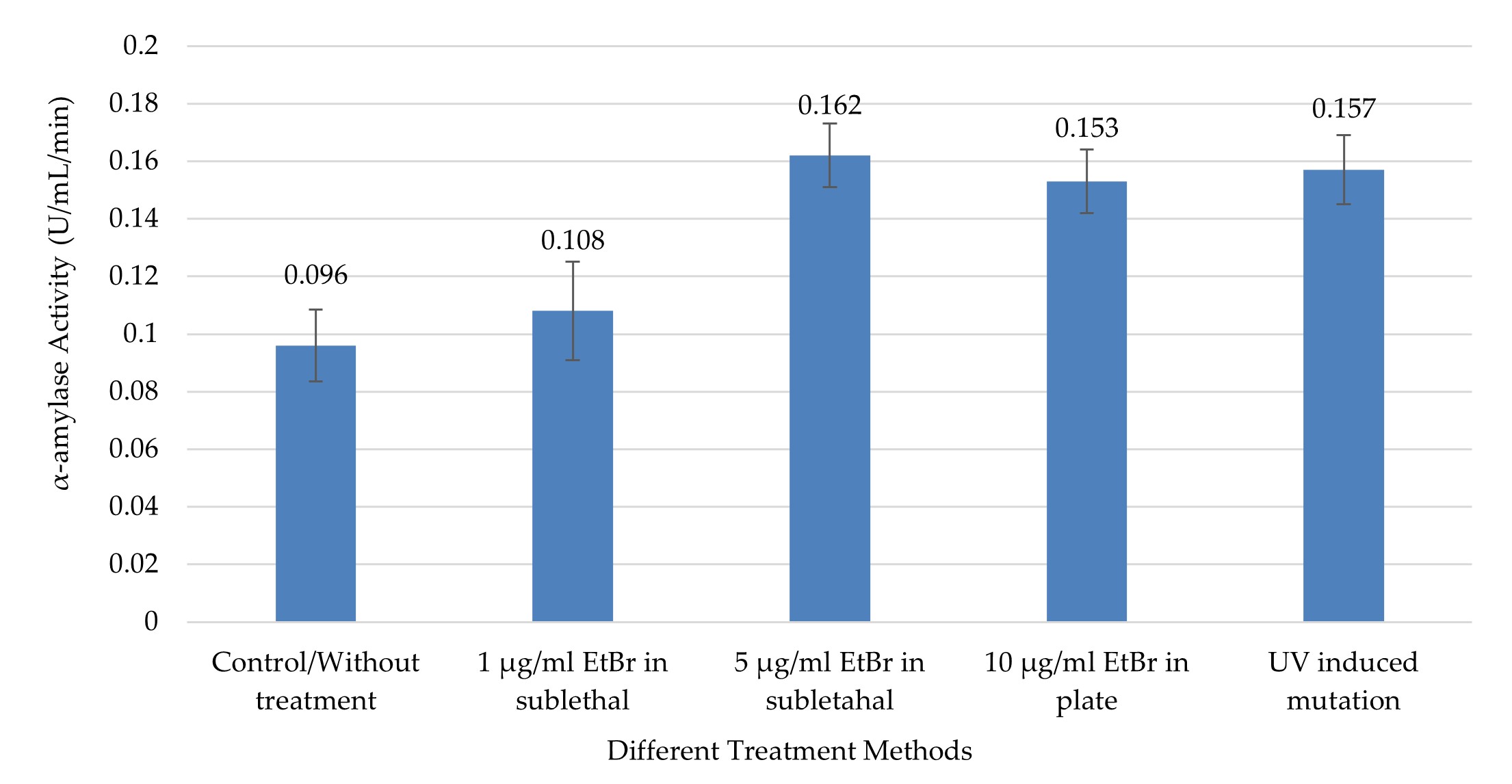{kind=link}
{kind=link}
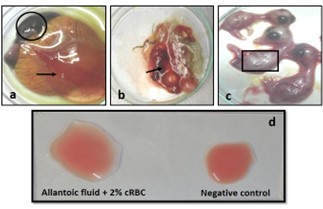{kind=link}
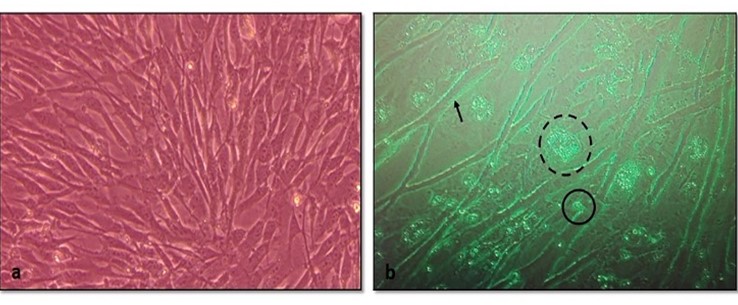{kind=link}
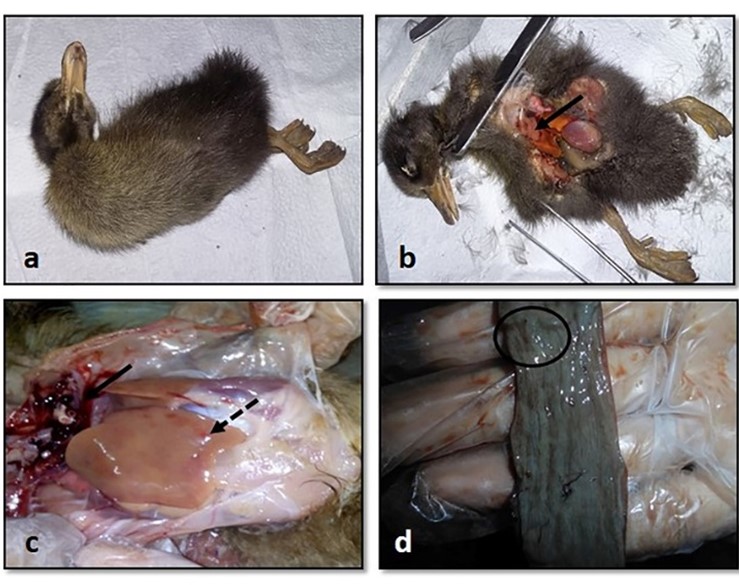{kind=link}
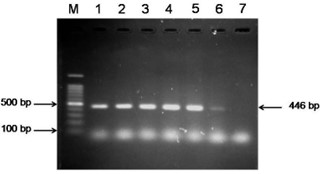{kind=link}
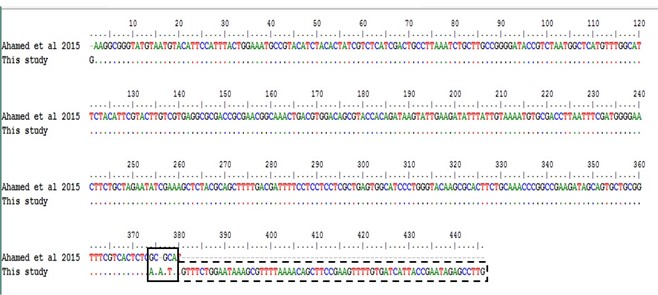{kind=link}
{kind=link}
{kind=link}
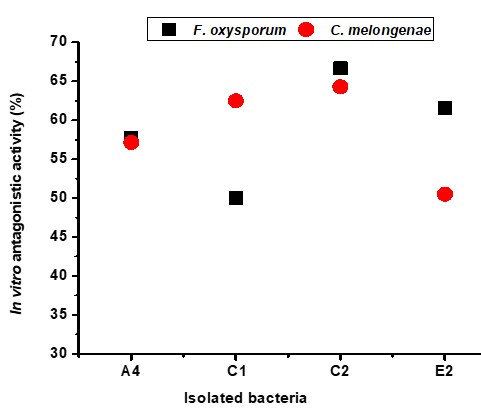{kind=link}
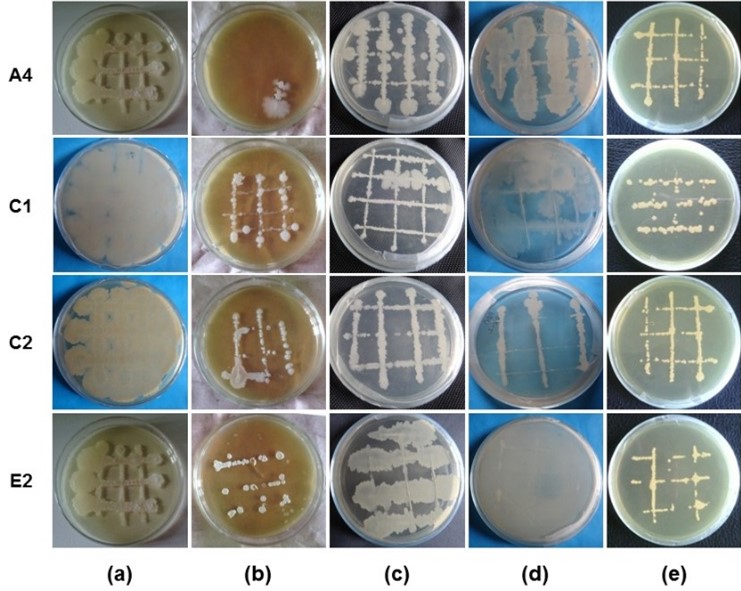{kind=link}
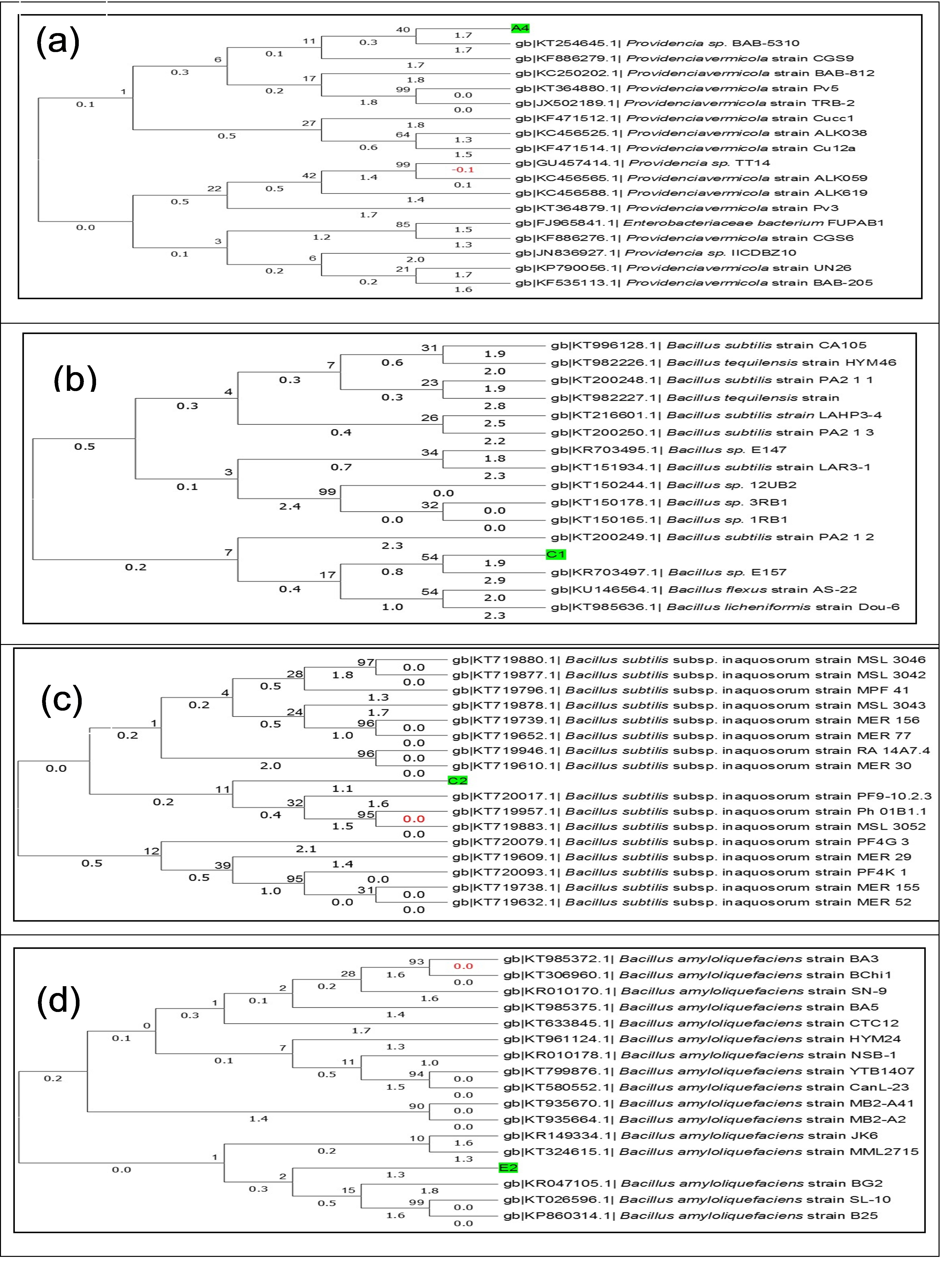{kind=link}
{kind=link}
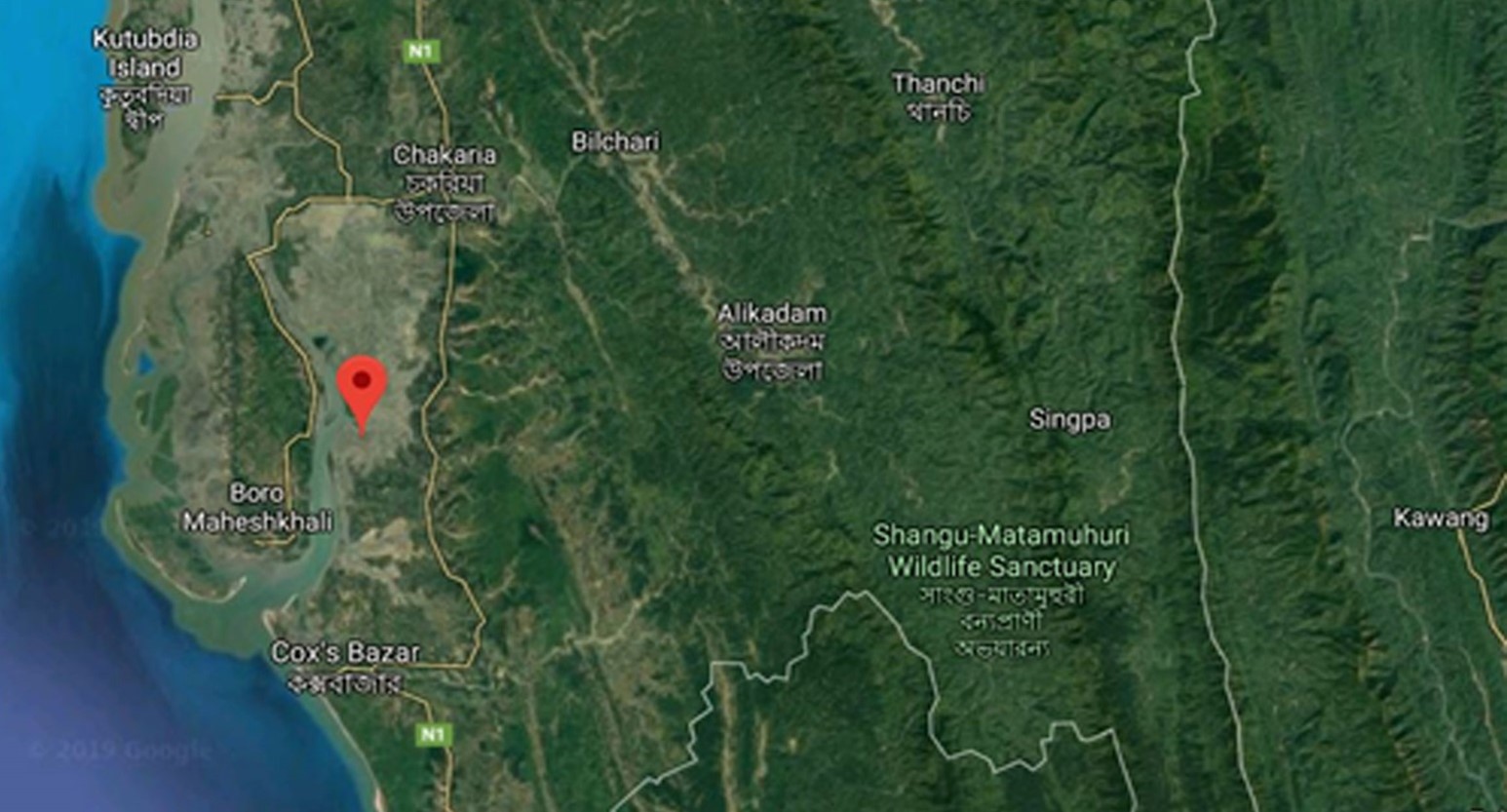{kind=link}
{kind=link}
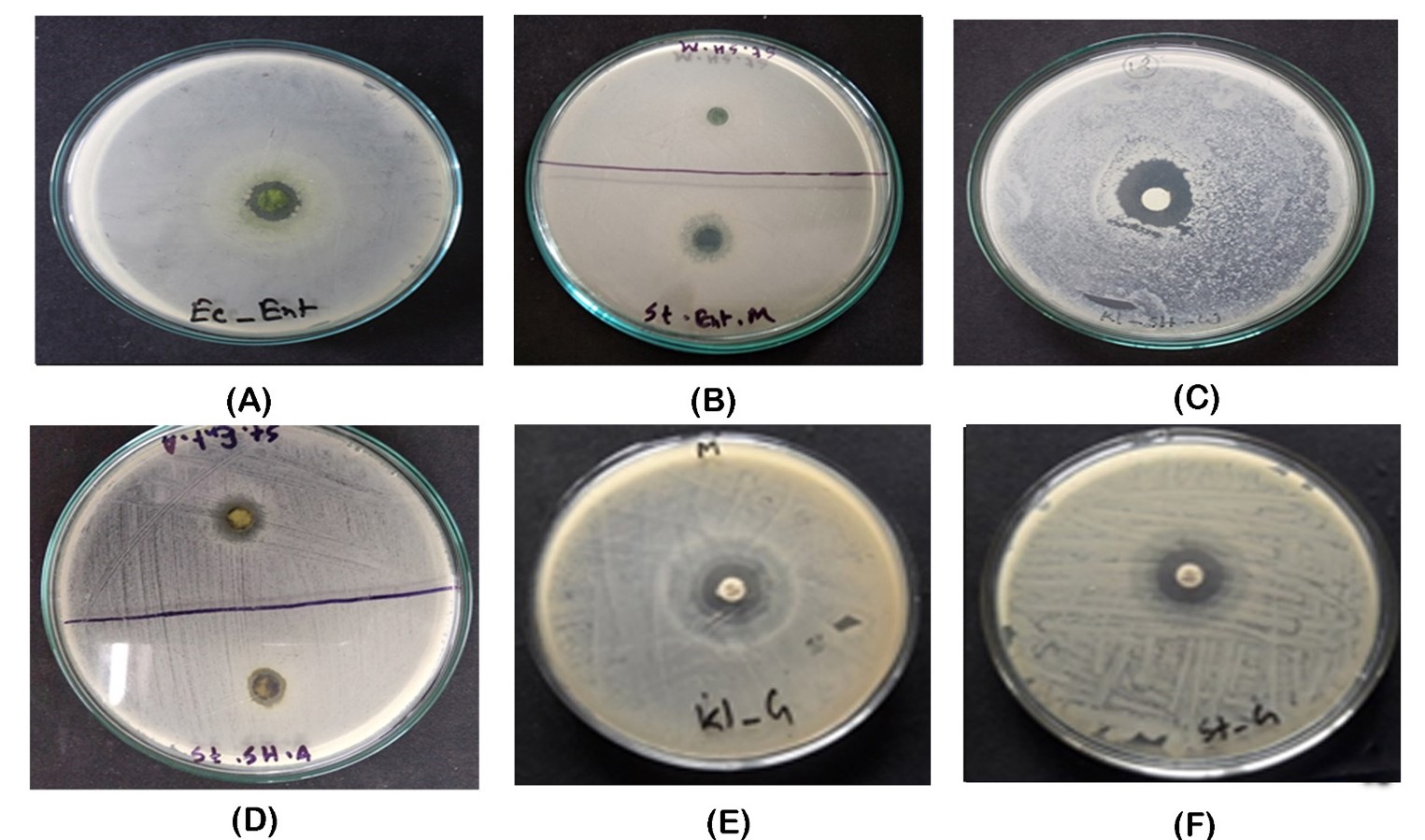{kind=link}
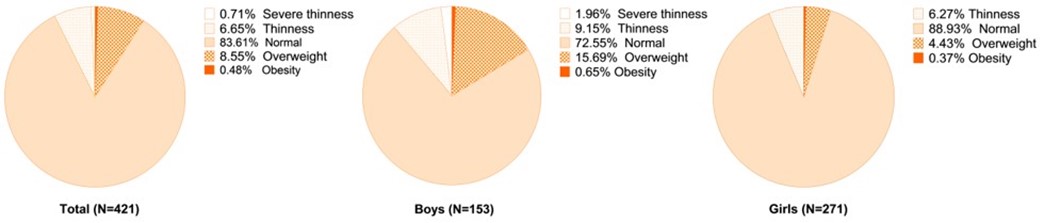{kind=link}
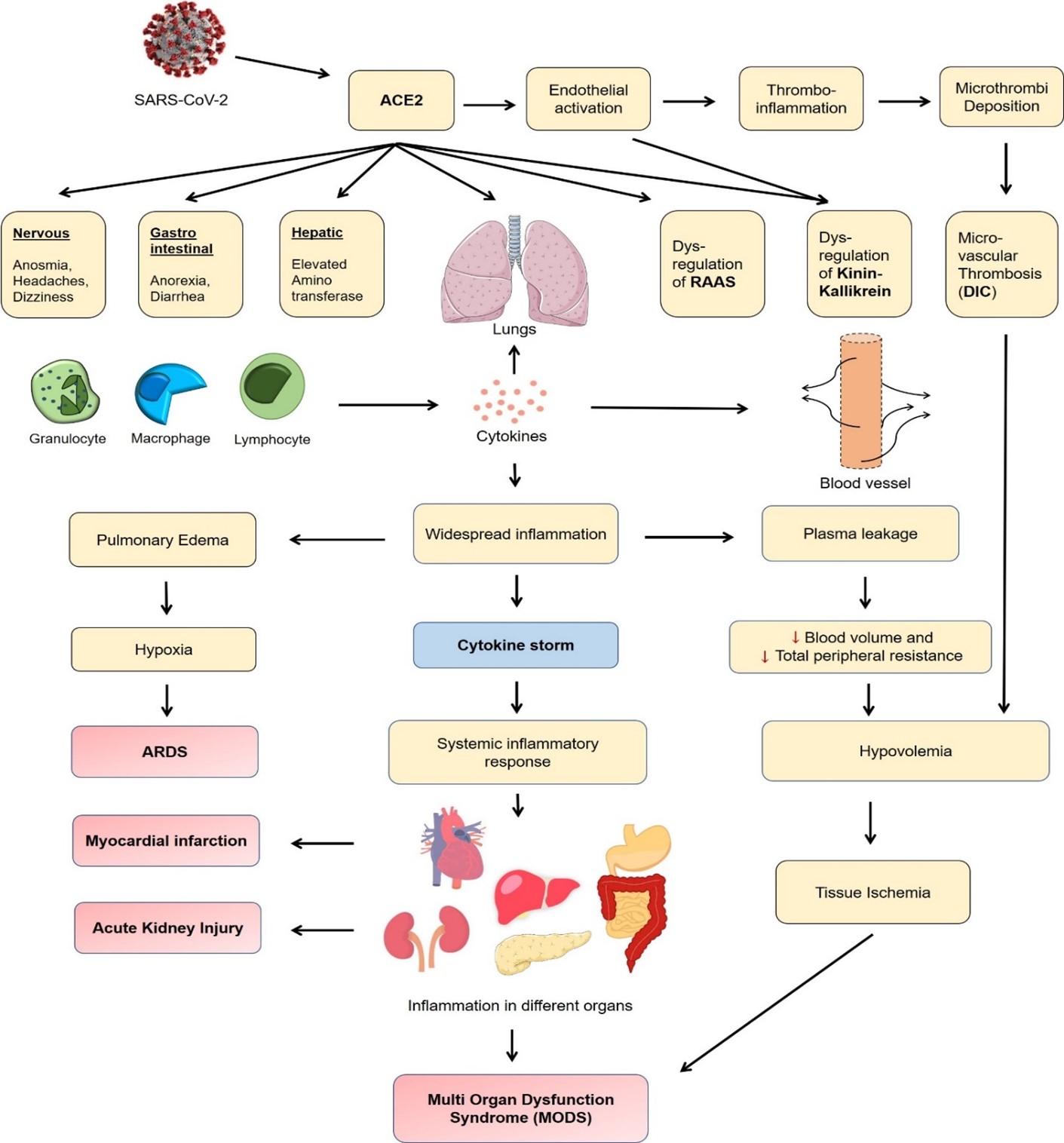{kind=link}
{kind=link}
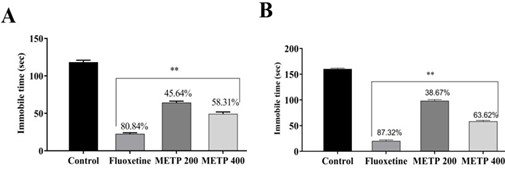{kind=link}
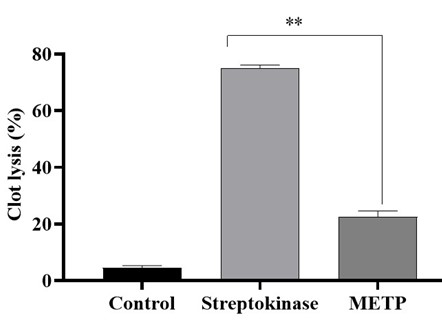{kind=link}
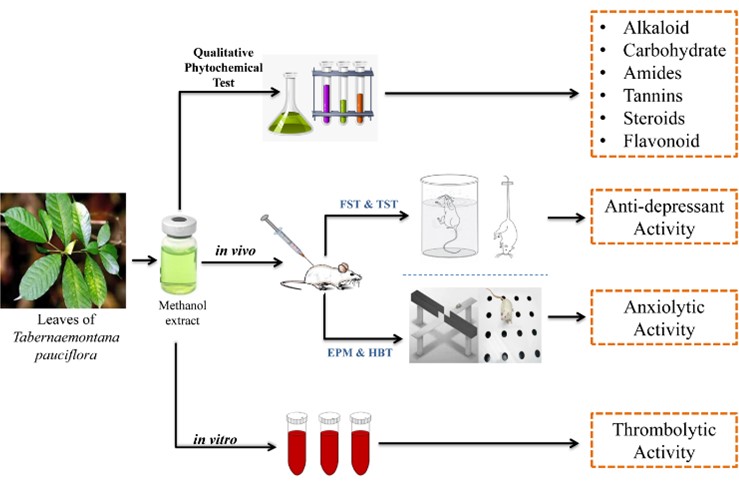{kind=link}
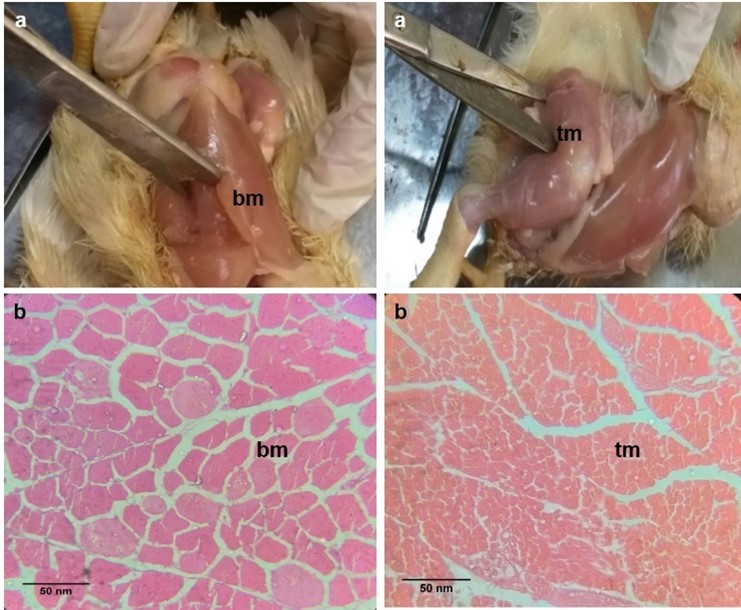{kind=link}
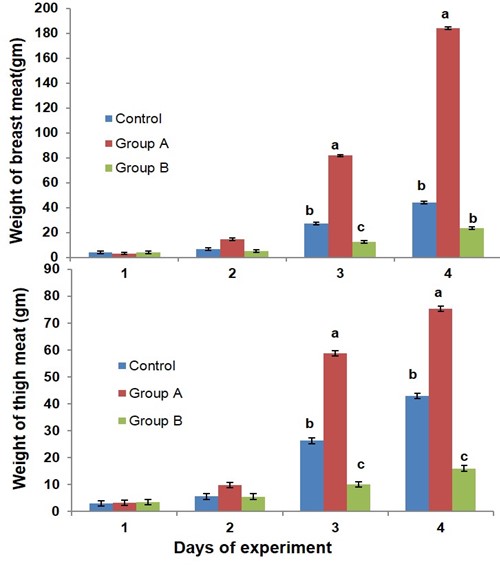{kind=link}
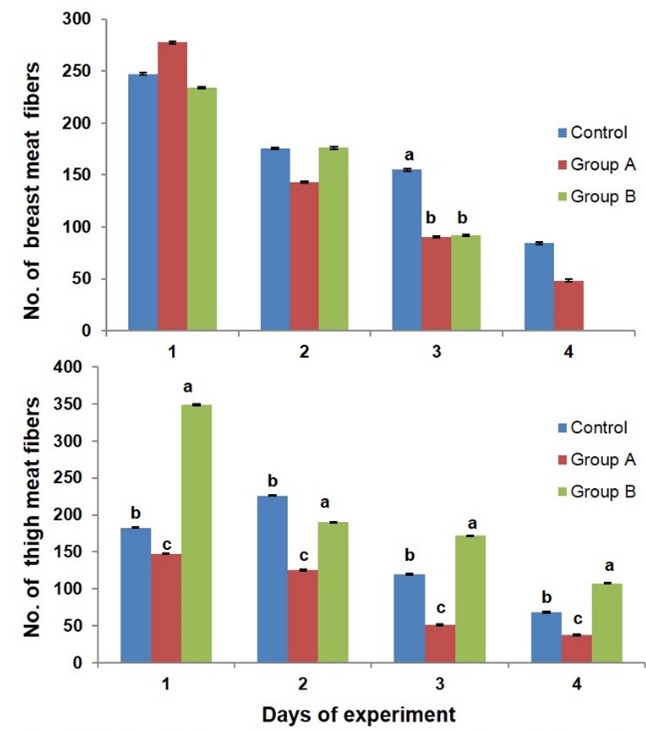{kind=link}
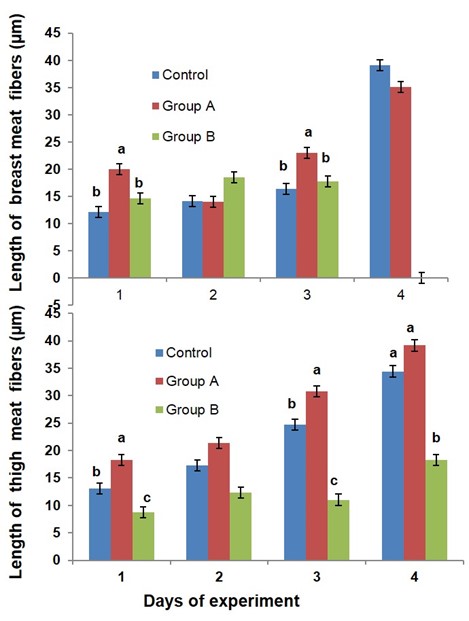{kind=link}
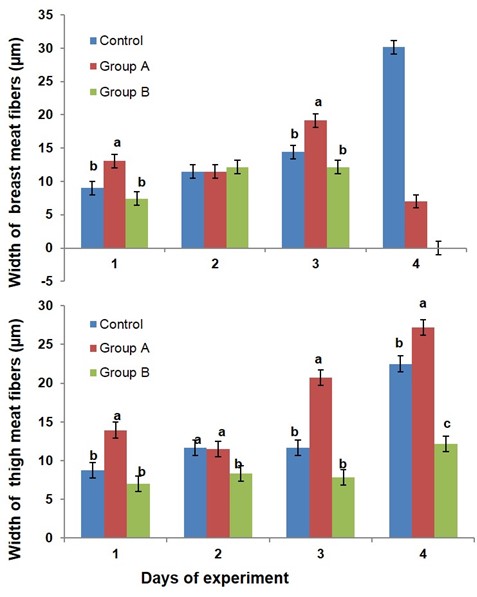{kind=link}
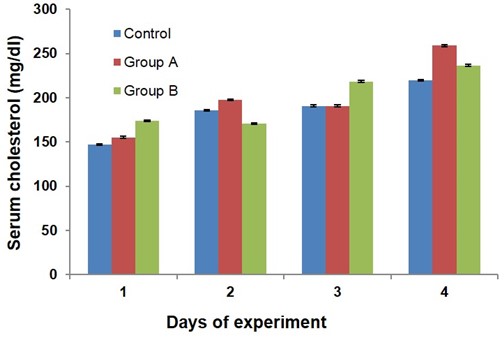{kind=link}
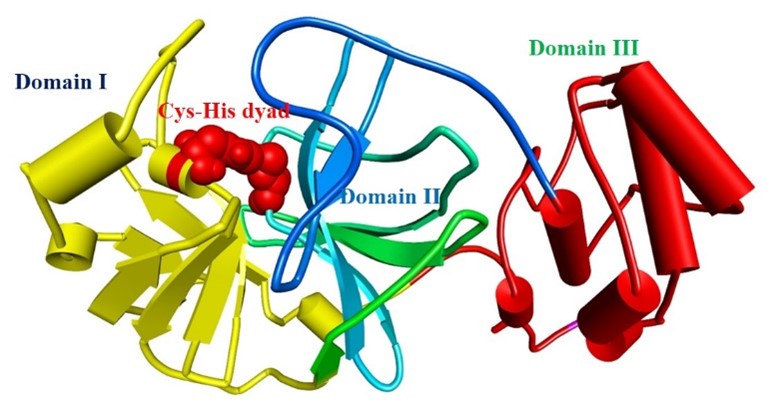{kind=link}
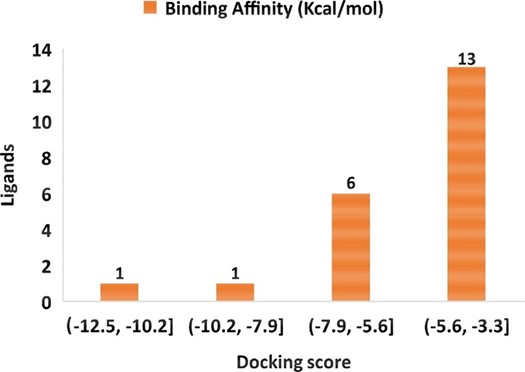{kind=link}
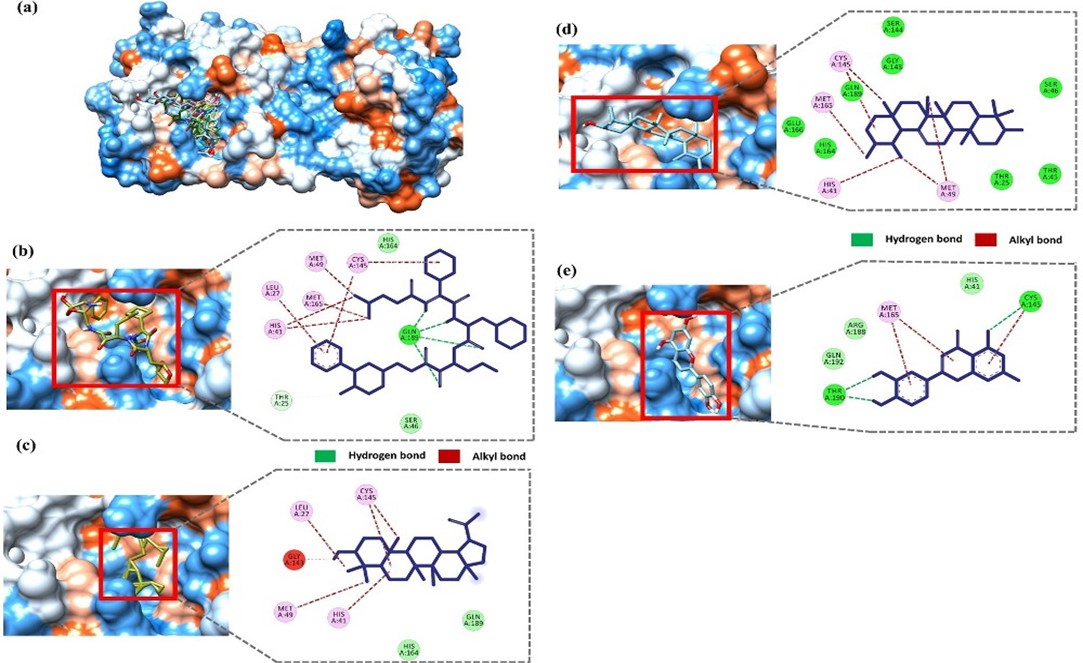{kind=link}
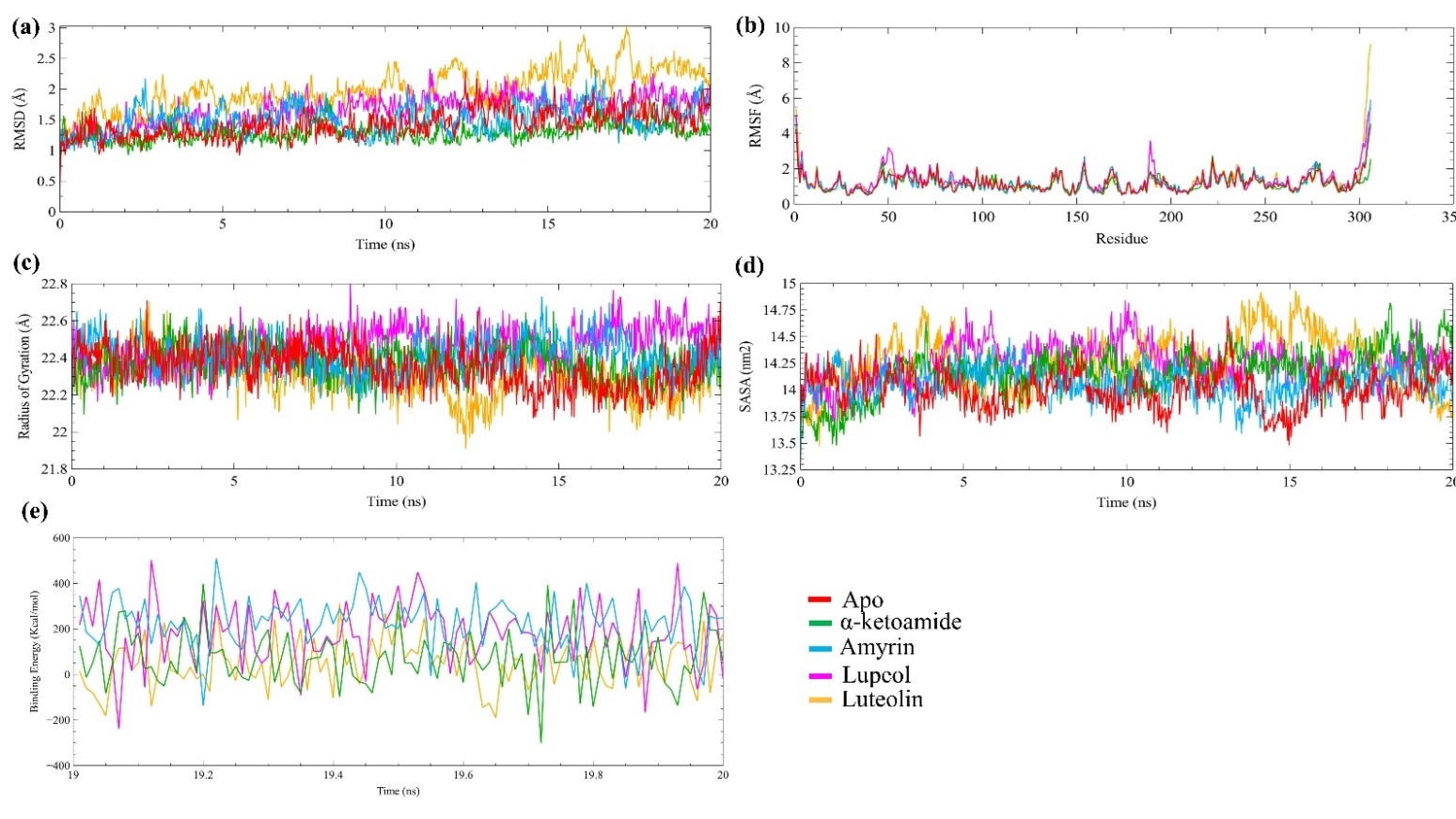{kind=link}
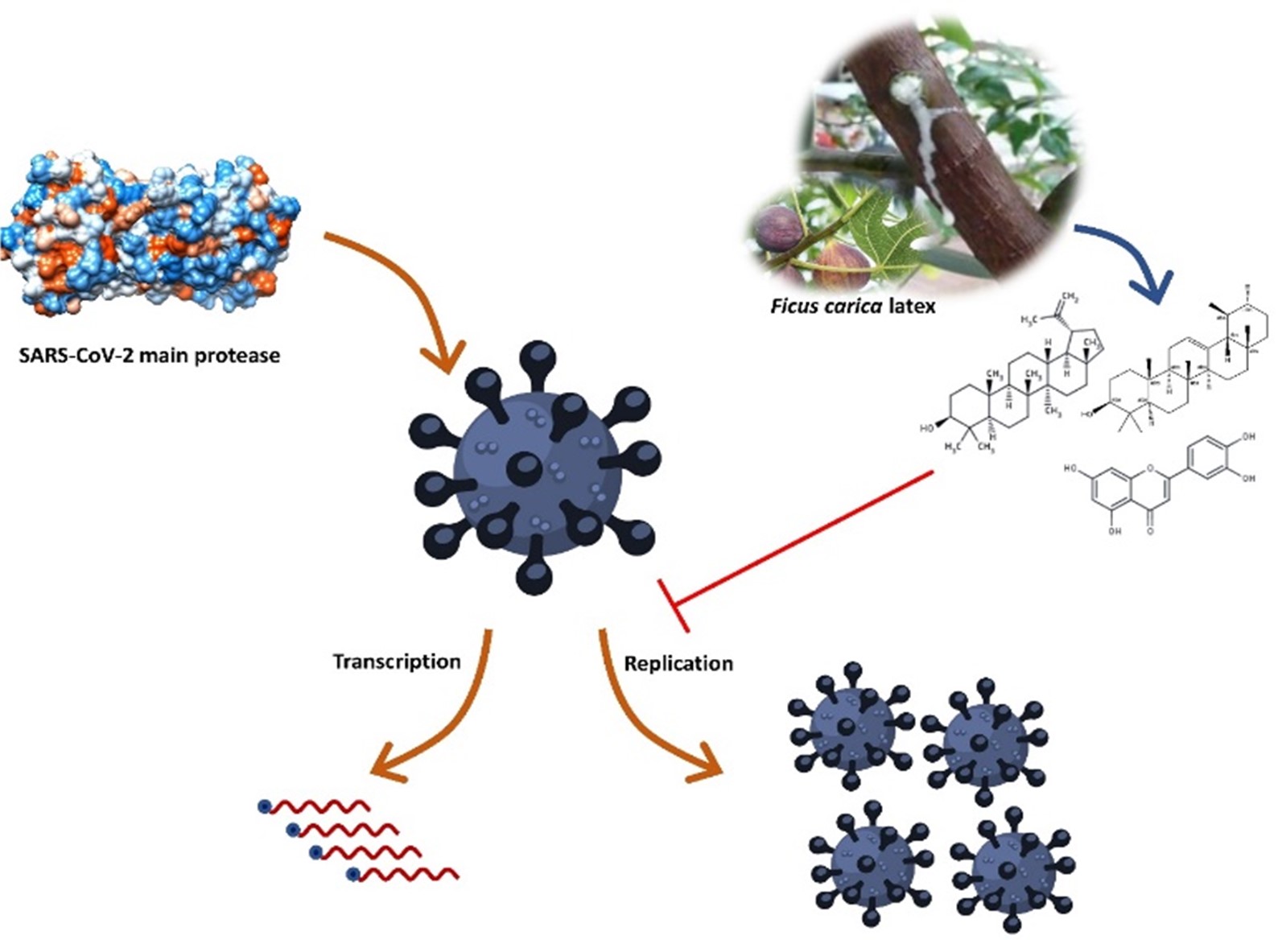{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}