HTTP 200 OK
Allow: GET, HEAD, OPTIONS
Content-Type: application/json
Vary: Accept
{
"count": 321,
"next": "https://jabet.bsmiab.org/articles/?format=api&page=17",
"previous": "https://jabet.bsmiab.org/articles/?format=api&page=15",
"results": [
{
"id": 132,
"slug": "178-1655373756-isolation-and-identification-of-salmonella-spp-and-escherichia-coli-from-water-used-during-live-transportation-of-pangasius-catfish-pangasianodon-hypophthalmus",
"featured": false,
"slider": false,
"issue": "Vol5 Issue3",
"type": "original_article",
"manuscript_id": "178-1655373756",
"recieved": "2022-06-16",
"revised": null,
"accepted": "2022-07-22",
"published": "2022-07-28",
"pdf_file": "https://jabet.bsmiab.org/media/pdf_file/2023/17/178-1655373756.pdf",
"title": "Isolation and identification of Salmonella spp. and Escherichia coli from water used during live transportation of Pangasius catfish, Pangasianodon hypophthalmus",
"abstract": "<p>Pangasius catfish (<em>Pangasianodon</em> <em>hypophthalmus</em>) is popular among fish farmers of Bangladesh due to its hardy characteristics, fast growth, and its ability to survive in high densities. Many consumers love to buy this fish, especially as live condition, due to its low market price and delicious fleshy meat. Bacterial outgrowth in transport water is frequent consequence including some enteric groups like <em>Salmonella </em>spp. and <em>E</em>. <em>coli</em>. The study attempted to know the occurrence of <em>Salmonella</em> spp. and <em>E</em>. <em>coli</em> in water used during live transportation of Pangasius catfish in Bangladesh. Water samples were collected from three different Pangasius catfish transportation channels at 2 h interval on-board transportation vehicle. The collected waters were then plated onto SS and EMB agar plates and 15, 20, and 17 suspected isolates were obtained from channel 1, 2 and 3, respectively. The isolates were confirmed through PCR techniques; <em>Salmonella</em> spp. was found only in channel 1 while <em>E</em>. <em>coli</em> were found in all 3 sampling channels under investigation. Among the suspected isolates, 13 isolates were positive for <em>E</em>. <em>coli</em> in channel 1, while 16 in both channel 2 and 3. Among the suspected isolates, 86.54% was <em>E. coli</em> positive, 1.92% was <em>Salmonella</em> positive, and 11.54% isolates were unidentified. The results indicated that the fishes were contaminated with <em>Salmonella </em>spp. and <em>E</em>. <em>coli</em> species either in the culture systems or during handling and live transportation.</p>",
"journal_reference": "J Adv Biotechnol Exp Ther. 2022; 5(3): 676-686.",
"academic_editor": "Md. Masudur Rahman, PhD; Sylhet Agricultural University, Bangladesh",
"cite_info": "Bhuiyan ANMRK, Hossain MM, et al. Isolation and identification of Salmonella spp. and Escherichia coli from water used during live transportation of Pangasius catfish, Pangasianodon hypophthalmus. J Adv Biotechnol Exp Ther. 2022; 5(3): 676-686.",
"keywords": [
"Salmonella spp.",
"Transport water",
"E. coli",
"Live transportation",
"Pangasius catfish"
],
"DOI": "10.5455/jabet.2022.d146",
"sections": [
{
"section_number": 1,
"section_title": "INTRODUCTION",
"body": "<p>Pangasius catfish (<em>Pangaisianodon hypophthalmus</em>; Sauvage, 1878) is a freshwater benthopelagic fish of the Pangasiidae family and has been recognized as a commercial aquaculture species in many Asian countries including Bangladesh. Currently, the species contributes 18.37% of total aquaculture production of Bangladesh [<a href=\"#r-1\">1</a>]. As Pangasius catfish is popular to consumers due to its low market value [<a href=\"#r-2\">2</a>], it is transported in live condition from production sites to markets in several areas of Bangladesh [<a href=\"#r-3\">3</a>]. Live transportation of Pangasius catfish includes harvesting it from the culture system, holding it in a confined plastic tank with water and carrying it to the desired retail market. Live transportation results in significant degradation of water quality parameters [<a href=\"#r-4\">4, 5</a>], thus it may cause deteriorative changes in the fishes. Bacteria grow in the transport water can even make the water quality worse for fish during live transportation. Previous reports showed that the viable bacterial counts of the transport water increased significantly with the periods of live transportation of Pangasius catfish [<a href=\"#r-3\">3</a>] and climbing perch, <em>Anabas testudineus </em>[<a href=\"#r-6\">6</a>] in Bangladesh.<br />\r\nBacteria of enteric origin (<em>Enterobacteriaceae</em>) are commonly disseminating from the gastrointestinal tract of humans and other animals are reported as common in aquatic environments [<a href=\"#r-7\">7, 8</a>], which may normally present in different parts of the apparently healthy fishes from contaminated waters. Among the <em>Enterobacteriaceae</em> species,<em> Salmonella</em>, <em>Escherichia coli</em>, and <em>Yarsenia enterocolitica</em> are not typically found in water or aquatic goods [<a href=\"#r-9\">9,10</a>]. <em>Salmonella</em> was not first identified in fish and is not a biological contaminant. It enters food through polluted water or inappropriate handling [<a href=\"#r-11\">11</a>], and both <em>Salmonella</em> and <em>E. coli</em> are regarded as public health hazards because they can cause food poisoning. The location, cultivated species, breeding procedures, processing, and cultural practices are the key elements that affect the risk of microbial contamination in aquaculture products. Some of these possible microbial risks could be brought on by subpar hygiene standards, sewage, and livestock drainage. Leaching, for instance, introduces environmental toxins into river waters, where they end up in the fish and have detrimental impacts on this ecosystem [<a href=\"#r-12\">12</a>]. In addition, massive use of fertilizer in the fish culture system and rearing fish with other types of animals, such as poultry, cattle and pigs are also responsible for contaminating the fish with <em>Salmonella</em> spp. [<a href=\"#r-13\">13-15</a>].<br />\r\nThe wide range of human diseases caused by <em>Salmonella </em>includes, enteric fever, bacteremia and gastroenteritis [<a href=\"#r-16\">16</a>]. <em>Salmonella </em>is a second leading cause of foodborne illness worldwide [<a href=\"#r-17\">17</a>]. The majority of human gastroenteritis by the <em>Salmonella</em> is caused through the ingestion of undercooked eggs, shellfish and fish [<a href=\"#r-18\">18</a>]. Different studies have been reported that freshwater fishes are contaminated by <em>Salmonella </em>spp. from the area where they were reared [<a href=\"#r-19\">19</a>]. <em>Salmonella</em> infections in freshwater fish are typically caused by faecal contamination of the water where the fish were caught [<a href=\"#r-19\">19</a>]. High prevalence of <em>Salmonella</em> in catfish was reported [<a href=\"#r-20\">20</a>]. The high temperature of the pond water, which enhances the organism’s growth rate, was accounted for the high prevalence rate [<a href=\"#r-21\">21</a>]. The most prevalent coliform in the intestinal flora of warm-blooded animals, however, is <em>E</em>. <em>coli</em>, which is assumed to be mostly related to faecal contamination [<a href=\"#r-22\">22</a>]. As a result, the potential that fish could serve as carriers of human pathogenic bacteria is receiving more attention [<a href=\"#r-23\">23-25</a>], as a wide range of bacteria including <em>Salmonella </em>spp. and <em>E. coli </em>have been isolated from skin, digestive tracts, kidney and muscle of different fish species of both temperate and tropical waters [<a href=\"#r-26\">26, 27</a>]. As common practice, underground water is used during live transportation of the fishes in Bangladesh [<a href=\"#r-28\">28</a>] and<em> Salmonella</em> spp. was not identified in the skin and muscle of Pangasius catfish from several ponds in Bangladesh, while it was isolated from the same species after marketing. On the other hand, <em>E</em>. <em>coli</em> was found in the skin and muscle of Pangasius catfish before and after marketing [<a href=\"#r-28\">28</a>]. Thus, it is not clear how the fishes are contaminated by these two pathogens of enteric origin. We also do not know whether the fishes are contaminated even during live transportation.<br />\r\nBased on the above background, the objective of this investigation was to assess the presence of <em>Salmonella </em>spp. and <em>E</em>. <em>coli</em> in water used during live transportation of the Pangasius catfish in Bangladesh. Investigations were done in three supply channels of Pangasius catfish and water samples were evaluated at 2 h intervals from the loading of the fishes.</p>"
},
{
"section_number": 2,
"section_title": "MATERIALS AND METHODS",
"body": "<p><strong>Descriptions of the studied supply channels of the Pangasius catfish</strong><br />\r\nThe study was conducted in three different Pangasius catfish supply channels of Bangladesh from July to December 2019. As Trishal Upazila of Mymensingh district is one of the major Pangasius catfish producing sites in Bangladesh, all the channels started from this Upazila. The supply channels were designated as “Channel 1” from Trishal, Mymensingh to Dhaka, “Channel 2” from Mymensingh to Faridpur, and “Channel 3<strong>”</strong> from Mymensingh to Sylhet (<a href=\"#figure1\">Figure 1</a>).</p>\r\n\r\n<div id=\"figure1\">\r\n<figure class=\"image\"><img alt=\"\" height=\"485\" src=\"/media/article_images/2023/10/25/178-1655373756-Figure1.jpg\" width=\"483\" />\r\n<figcaption><strong>Figure 1. </strong>Map showing the location of sample supply channels of live Pangasius catfish (<em>Pangasianodon hypophthalmus</em>) transportation in Bangladesh. Channel-1: Trishal, Mymensingh to Kawran Bazar, Dhaka; Channel-2: Trishal, Mymensingh to Faridpur; and Channel-3: Trishal, Mymensingh to Poschim Kazir Bazar, Sylhet. The map is extracted from DIVA-GIS using Geographical Information System (GIS) and visualized by ArcMap version 10.7.</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Harvesting and preparation before live transportation</strong><br />\r\nPangasius catfish were harvested with the surrounding net and prepared for transportation at dusk. A very brief conditioning period was allowed prior to transportation. In most cases, transportation started at night and reached to the final destination/ unloading points (retail markets) at dawn. Around 40 kg fishes (20-22 fishes) were loaded in plastic made transportation tank having a capacity of 1000 liters. But the tanks were filled half with 500 liters of deep tube-well water. Approximately 40-42 transportation tanks were incorporated in a commercial vehicle (Truck). Exchange of water of transportation tank with the deep tube-well water once after 2-3 hours during live transportation of Pangasius catfish was commonly practiced in all the supply channels during transportation. At the retail market, Pangasius catfish were collected from the transportation tank and live fishes were separated from the dead ones.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Isolation of <em>Salmonella</em> spp. and <em>E.</em> <em>coli</em></strong><br />\r\nThe isolation and identification of <em>Salmonella </em>spp. and <em>E</em>. <em>coli</em> was carried out based on culture on SS and EMB agar plates with slight modifications as described [<a href=\"#r-29\">29</a>]. Plates with SS agar and EMB agar (HIMEDIA, India) were prepared and used for the isolation of enteric bacteria from water used during live transportation of Pangasius catfish. Water samples were collected from the Pangasius catfish transportation tank with previously sterilized plastic bottle from the start of transportation (loading) to the end (unloading) at every 2 h interval. After collection, water samples were spread on previously prepared SS and EMB agar plates immediately. The plates were brought to the Laboratory of Fisheries Microbiology, Department of Fisheries Technology, Bangladesh Agricultural University, Mymensingh, Bangladesh and incubated at 37 for 24 h. After incubation, colourless or translucent, black, pink-coloured colonies were observed on SS agar. Suspected colonies on EMB agar had black or dark centre with or without green metallic sheen. The resulted colonies from SS and EMB agar plates were subjected to subcultures again on SS and EMB agar plates in order to obtain isolated colonies for pure culture. The resulted isolated colonies were kept on previously prepared agar slant of plate count agar (HIMEDIA, India) for further analysis. The preserved colonies were then streaked on SS agar plate from the agar slant and the plates were incubated at 37 for 24 h. After incubation, the resulted colonies were used for identification of <em>Salmonella</em> spp. and <em>E.</em> <em>coli</em>.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Identification of <em>Salmonella</em> spp. and <em>E.</em> <em>coli</em></strong><br />\r\nPreviously obtained isolates were incubated in nutrient broth for 24 h at 37 in order to increase the number of bacteria to extract DNA. For this purpose, a pure colony was transferred in a test tube containing 10 ml of previously prepared nutrient agar broth. The broth was then incubated at 37 for 24 h. After incubation, turbidity of the nutrient broth ensured the growth of bacteria. Then 1 ml of nutrient broth was taken in a sterilized eppendorf and centrifuged at 5,000 rpm for 3 min. The settlings on the eppendorf were mixed with 100 μl distilled water and again centrifuged at 5,000 rpm for 3 min. After that, the settlings were mixed with 100 μl distilled water and homogenized and subjected to boiling for 10 min followed by keeping in ice for cold shock. Again, centrifugation was done at 10,000 rpm for 10 min. Finally, the supernatant was collected and used as DNA template for PCR.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>PCR amplification of extracted bacterial DNA</strong><br />\r\nTwo sets of pre-tested primers were used for identification of <em>Salmonella</em> spp. and <em>E. coli</em> (<a href=\"#Table-1\">Table 1</a>). For <em>Salmonella</em> spp., <em>invA</em> gene was targeted, and used forward primer, <em>InvA </em>F (ATCAGTACCAGTCGTCTTATCTTGAT) and reverse primer, <em>InvA </em>R (TCTGTTTACCGGGCATACCAT) for amplification [<a href=\"#r-27\">27</a>]. While <em>16S</em> rRNA gene was partially amplified using EC-1 (GACCTCGGTTTAGTTCACAGA) as forward primer and EC-2 (CACAGCTGACGCTGACCA) as reverse primer [<a href=\"#r-30\">30</a>]. For PCR amplification, 12.5 μl of master mix, 8.5 μl of nuclease free DEPC treated dH<sub>2</sub>O, 1 μl forward primer, 1 μl reverse primer and 2 μl of extracted bacterial genomic DNA were taken into the PCR tubes. The tubes were placed into the thermal cycler (2720 Thermal Cycler, Applied Biosystems, Waltham, USA) immediately after adding the master mix with the DNA, and the cyclic program was resumed after the program was over. PCR products were visualization by agarose gel-electrophoresis.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Agarose gel electrophoresis of PCR product</strong><br />\r\nFor electrophoresis, 1.5% agarose (Sigma-Aldrich, USA) gel was used for electrophoresis of the PCR products. Gel casting tray was assembled with gel comb of appropriate teeth size and number. 1.5% agarose solution was prepared in TAE buffer by melting in a microwave oven. Melted agarose was poured onto the casting tray and allowed to solidify on the bench. The hardened gel in its tray was transferred to the electrophoresis tank containing sufficient TAE buffer to cover the gel. The comb was gently removed. Then, 5 μl of each PCR product was mixed with 1 μl loading buffer and the sample was loaded to the appropriate well of the gel. In addition, 5 μl DNA size marker was loaded in one well. The leads of the electrophoresis apparatus were connected to the power supply and the electrophoresis was run at 100V. When DNA migrated sufficiently as judged from the migration of bromophenicol blue of loading buffer, the power supply was disconnected. The gel was stained in ethidium bromide (0.5 μg/ml) for 10 min in a dark place. Then the gel was de-stained in distilled water for 10 min. De-stained gel was then placed on the UV transilluminator (Biometra, Germany) in the dark chamber of the image documentation system. The UV light of the system was switched on and the image was viewed on the monitor, focused, acquired, and saved in an USB flash drive.</p>"
},
{
"section_number": 3,
"section_title": "RESULTS",
"body": "<p><strong>Isolation of <em>Salmonella </em>spp. and <em>E</em>. <em>coli</em></strong><br />\r\nIn order to detect <em>Salmonella </em>spp. and <em>E</em>. <em>coli</em>, water used in live transportation of Pangasius catfish was collected from three marketing channels. The collected water samples were then inoculated onto SS and EMB agar plates and well-separated colonies were obtained, regarded as isolates. A total of 52 isolates were collected depending on the characteristics of the colony on SS and EMB agar plates.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Identification of <em>Salmonella </em>spp. and <em>E</em>. <em>coli</em></strong><br />\r\nThe DNA of all the isolates were extracted and subjected to PCR according to the protocol described in the methodology section. Predefined <em>invA</em> genes of <em>Salmonella </em>spp. and parts of 16S rRNA genes of <em>E</em>. <em>coli</em> were targeted using two different sets of primers namely, <em>InvA</em> (211 bp) and ECO (585 bp) for <em>Salmonella </em>spp. and <em>E</em>. <em>coli,</em> respectively (<a href=\"#Table-1\">Table 1</a>). <em>Salmonella </em>spp. positive was confirmed considering 211 bp bands on gel after electrophoresis. For <em>E</em>. <em>coli, </em>bands of 585 bp on gel were considered positive (<a href=\"#figure2\">Figure 2</a> and <a href=\"#figure3\">3</a>).</p>\r\n\r\n<div id=\"Table-1\">\r\n<p><a href=\"https://jabet.bsmiab.org/table/178-1655373756-table1/\">Table-1</a><strong>Table 1. </strong>PCR protocol used for the confirmation of <em>Salmonella </em>spp. and <em>E</em>. <em>coli</em>.</p>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<div id=\"figure2\">\r\n<figure class=\"image\"><img alt=\"\" height=\"245\" src=\"/media/article_images/2023/10/25/178-1655373756-Figure2.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 2.</strong> Percentage of <em>Salmonella</em> spp. and <em>E. coli</em> positive. The isolates were obtained from water used during live transportation of Pangasius catfish (<em>Pangasianodon hypophthalmus</em>) in Bangladesh. The arrow showing the amplicon size (211 bp), lane L = 100 bp DNA ladder, lane P = positive control, lane N = negative control, and Lane 1 = positive results (only one isolate was found positive among 52).</figcaption>\r\n</figure>\r\n</div>\r\n\r\n<div id=\"figure3\">\r\n<figure class=\"image\"><img alt=\"\" height=\"275\" src=\"/media/article_images/2023/10/25/178-1655373756-Figure3.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 3. </strong>Results of agarose gel electrophoresis of <em>16S rRNA </em>gene for <em>E. coli </em>confirmation. The isolates were obtained from water used during live transportation of Pangasius catfish (<em>Pangasianodon hypophthalmus</em>) in Bangladesh. The arrow showing the amplicon size (585 bp), lane L = 100 bp DNA ladder, lane P = positive control, and Lane 1-52 = isolates tested for <em>E. coli </em>(45 isolates were found positive among 52).</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Incidence of <em>Salmonella </em>spp. and <em>E</em>. <em>coli</em> in water used during live transportation of Pangasius catfish</strong><br />\r\nOut of 15 isolates from sampling Channel 1, only 1 isolate (6.67%) was positive for <em>Salmonella </em>spp., while 13 isolates (86.66%) were positive for <em>E</em>. <em>coli</em> and 1 isolate (6.67%) remained unidentified. In Channel 2, 16 <em>E</em>. <em>coli </em>positive isolates were identified which accounts for 80% of the total collected isolates and 4 isolates (20%) remained unidentified. In sampling Channel 3 from 17 isolates, 16 isolates (94.12%) were identified as positive for <em>E</em>. <em>coli</em> and 1 isolate (5.88%) remained unidentified. On the other hand, 45 isolates (86.54%) were identified as positive for <em>E</em>. <em>coli</em> and the remaining 6 isolates (11.54%) were not confirmed as <em>Salmonella </em>spp. or <em>E</em>. <em>coli</em>, gave no band on electrophoresis (<a href=\"#Table-2\">Table 2</a>, and <a href=\"#figure4\">Figure 4</a>). Although, <em>E</em>. <em>coli</em> positive isolates were found in all the three sampling channels, only one <em>Salmonella </em>spp. positive isolate was confirmed obtained from the sampling Channel 1 (<a href=\"#Table-2\">Table 2</a> and <a href=\"#figure5\">Figure 5</a>).<br />\r\nNumber of isolates obtained from water used during live transportation of Pangasius catfish (<em>Pangasianodon hypophthalmus</em>) in Bangladesh. The water samples were collected from 3 different supply channels.</p>\r\n\r\n<div id=\"Table-2\">\r\n<p><a href=\"https://jabet.bsmiab.org/table/178-1655373756-table2/\">Table2</a><strong>Table 2. </strong>Positive isolates for <em>Salmonella </em>spp. and <em>E</em>. <em>coli.</em></p>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<div id=\"figure4\">\r\n<figure class=\"image\"><img alt=\"\" height=\"274\" src=\"/media/article_images/2023/10/25/178-1655373756-Figure4.jpg\" width=\"454\" />\r\n<figcaption><strong>Figure 4.</strong> Number of <em>Salmonella</em> spp., and <em>E. coli</em> positive isolates considering the total number of isolates obtained from water used during live transportation of Pangasius catfish (<em>Pangasianodon hypophthalmus</em>) in Bangladesh. The water samples were collected from 3 different supply channels.</figcaption>\r\n</figure>\r\n</div>\r\n\r\n<div id=\"figure5\">\r\n<figure class=\"image\"><img alt=\"\" height=\"334\" src=\"/media/article_images/2023/10/25/178-1655373756-Figure5.jpg\" width=\"454\" />\r\n<figcaption><strong>Figure 5. </strong>Percentage of <em>Salmonella</em> spp. and <em>E. coli </em>positive isolates obtained from water used during live transportation of Pangasius catfish (<em>Pangasianodon hypophthalmus</em>) in Bangladesh. The water samples were collected from 3 different supply channels.</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>"
},
{
"section_number": 4,
"section_title": "DISCUSSION",
"body": "<p>A previous study showed the presence of both <em>Salmonella</em> spp. and <em>E</em>. <em>coli</em> in Pangasius catfish collected from several retail markets regardless of their presence or absence in the same specimen before marketing. So, the experiment was designed to identifying the presence of <em>Salmonella</em> spp. and <em>E</em>. <em>coli</em> in water which was used in live transportation of Pangasius catfish from farms to retail markets. In order to achieve the objectives of the study, water samples were collected from the plastic barrels where Pangasius catfish were kept alive during transportation from farm to the retail markets at every 2 h interval from 3 different Pangasius catfish marketing channels of Bangladesh. We assumed that these two pathogens would be detected from the water subsamples collected from the barrels used in live transportation. The detection of <em>Salmonella </em>spp. and <em>E</em>. <em>coli</em> from the water used in live transportation of Pangasius catfish is of great concern due to its potential to cause enteric disease [<a href=\"#r-31\">31</a>]. The water samples were taken aseptically from the plastic barrels where Pangasius catfish were carried during transportation. After collection, water samples were then placed on SS and EMB agar plates at the start of transportation to the end at 2 h interval. Upon completion of sampling, plates were brought to the laboratory and incubated at 37 for 24 h. After growth of microorganisms on SS and EMB agar plates, suspected <em>Salmonella</em> spp. and <em>E</em>. <em>coli</em> colonies were isolated according to a protocol with slight modifications as described [<a href=\"#r-29\">29</a>]. The collected isolates of <em>Salmonella</em> spp. and <em>E</em>. <em>coli</em> from subsamples water were identified through PCR using two primers which are very fast, unique and sensitive in identifying the target gene by PCR technique [<a href=\"#r-32\">32</a>].<br />\r\nHighest incidence of <em>E</em>. <em>coli</em> was found in sampling Channel 3 and the lowest was in sampling Channel 2. Incidence of <em>Salmonella</em> spp. was seen only in the sampling Channel 1. Among the collected isolates regardless of the sampling channels, 86.54% were identified as positive for <em>E</em>. <em>coli</em> whereas only 1.92% <em>Salmonella </em>spp. positive isolates were identified. On the other hand, 11.54% isolates remained unidentified. In comparison to <em>Salmonella </em>spp., percentages of <em>E</em>. <em>coli</em> positive isolates were always higher in the studied channels. Fish act as carrier of microorganisms as it is continuously exposed to the microorganisms in aquatic environment and the microorganism in fish reflects the conditions of the environment. At the time of harvesting, a wide variety of microorganisms contain in the body of fresh fish which are known as the microflora of that fish [<a href=\"#r-33\">33</a>]. Fish or processed fish may be contaminated with different types of bacteria, such as <em>Salmonella</em>, coliform, faecal coliform, <em>Streptococcus</em>, <em>Staphylococcus aureus</em> and these are responsible for different types of foodborne disease [<a href=\"#r-34\">34</a>]. In our experiment, <em>Salmonella</em> spp. was found only in sampling Channel 1, while <em>E</em>. <em>coli</em> was identified from all the sampling channels. Identification of these enteric bacteria indicated that the water was polluted with faecal matters. Bird droppings, human faeces and other animal faeces falling into aquatic environment are contaminated with enteric bacteria. Deep tube well water was used during live transportation of Pangasius catfish. In addition, stated that ground water was free from any sorts of coliform organisms, and they isolated <em>E</em>. c<em>oli </em>from the pond sediments and stated the sources as pigeon [<a href=\"#r-35\">35</a>]. Moreover, a study found <em>E</em>. <em>coli </em>in all the tube well water samples in their study while <em>Salmonella </em>spp. was absent in all tube well water samples [<a href=\"#r-26\">26</a>]. On the other hand, low concentration of faecal coliform bacteria in tube well water of Bangladesh [<a href=\"#r-36\">36</a>]. Groundwater can also be contaminated by a wide range of pathogens [<a href=\"#r-15\">15</a>, <a href=\"#r-37\">37-38</a>] and found <em>Salmonella </em>spp., and <em>E</em>. <em>coli</em> in ground water sample. Identified enteric bacteria would be the reflection of the ponds from where the Pangasius catfish were harvested. The presence of enteric bacteria in water, sediment and fish reflects the water used in transportation are transmitted by fish. Moreover, Unhygienic handling of Pangasius catfish during transportation may contaminate the fish with <em>Salmonella </em>spp. and <em>E. coli</em>. [<a href=\"#r-28\">28</a>]. It also stated that rough handling and lack of proper sanitation during transportation causes Pangasius and other fishes more contaminated collected from retail market compared to pond samples of the same species.</p>"
},
{
"section_number": 5,
"section_title": "CONCLUSIONS",
"body": "<p>Bacterial hazards are closely related to food safety. The prevalence of <em>Salmonella</em> spp. was lower in Pangasius catfish transportation water in comparison to <em>E</em>. <em>coli</em>. The presence of <em>Salmonella </em>spp., and <em>E</em>. <em>coli</em> in water used during live transportation of Pangasius catfish indicated that the fishes were harvested from water which was polluted with faeces. Water used in the transportation would be another source of enteric bacteria. Personnel involved in handling of fish during live transportation would be a potential source of enteric bacteria. These enteric microorganisms could cause foodborne illness among the consumers which ultimately affects the profit margin of farmers. So, it is necessary to maintain proper sanitation and handling during live transportation of Pangasius catfish to ensure food safety.</p>"
},
{
"section_number": 6,
"section_title": "ACKNOWLEDGEMENTS",
"body": "<p>The authors are grateful to the Department of Fisheries Technology, Bangladesh Agricultural University, for the laboratory and technical support and to the Bangladesh Agricultural University Research System (BAURES) for the research grants.</p>"
},
{
"section_number": 7,
"section_title": "AUTHOR CONTRIBUTIONS",
"body": "<p>The work was designed and supervised by MNH and MNU. The research work was performed by ANMRKB and MMH. The first draft of this manuscript was prepared by ANMRKB. MMH analyzed the data and improved the overview of the manuscript. MNH critically revised, improved and approved the final version of the manuscript.</p>"
},
{
"section_number": 8,
"section_title": "CONFLICTS OF INTEREST",
"body": "<p>There is no conflict of interest among the authors.</p>"
}
],
"figures": [
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/10/25/178-1655373756-Figure1.jpg",
"caption": "Figure 1. Map showing the location of sample supply channels of live Pangasius catfish (Pangasianodon hypophthalmus) transportation in Bangladesh. Channel-1: Trishal, Mymensingh to Kawran Bazar, Dhaka; Channel-2: Trishal, Mymensingh to Faridpur; and Channel-3: Trishal, Mymensingh to Poschim Kazir Bazar, Sylhet. The map is extracted from DIVA-GIS using Geographical Information System (GIS) and visualized by ArcMap version 10.7.",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/10/25/178-1655373756-Figure2.jpg",
"caption": "Figure 2. Percentage of Salmonella spp. and E. coli positive. The isolates were obtained from water used during live transportation of Pangasius catfish (Pangasianodon hypophthalmus) in Bangladesh. The arrow showing the amplicon size (211 bp), lane L = 100 bp DNA ladder, lane P = positive control, lane N = negative control, and Lane 1 = positive results (only one isolate was found positive among 52).",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/10/25/178-1655373756-Figure3.jpg",
"caption": "Figure 3. Results of agarose gel electrophoresis of 16S rRNA gene for E. coli confirmation. The isolates were obtained from water used during live transportation of Pangasius catfish (Pangasianodon hypophthalmus) in Bangladesh. The arrow showing the amplicon size (585 bp), lane L = 100 bp DNA ladder, lane P = positive control, and Lane 1-52 = isolates tested for E. coli (45 isolates were found positive among 52).",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/10/25/178-1655373756-Figure4.jpg",
"caption": "Figure 4. Number of Salmonella spp., and E. coli positive isolates considering the total number of isolates obtained from water used during live transportation of Pangasius catfish (Pangasianodon hypophthalmus) in Bangladesh. The water samples were collected from 3 different supply channels.",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/10/25/178-1655373756-Figure5.jpg",
"caption": "Figure 5. Percentage of Salmonella spp. and E. coli positive isolates obtained from water used during live transportation of Pangasius catfish (Pangasianodon hypophthalmus) in Bangladesh. The water samples were collected from 3 different supply channels.",
"featured": false
}
],
"authors": [
{
"id": 538,
"affiliation": [
{
"affiliation": "Department of Fisheries Technology, Bangladesh Agricultural University, Mymensingh-2202, Bangladesh"
}
],
"first_name": "A N M Rezvi Kaysar",
"family_name": "Bhuiyan",
"email": null,
"author_order": 1,
"ORCID": "http://orcid.org/0000-0001-7911-6871",
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 132
},
{
"id": 539,
"affiliation": [
{
"affiliation": "Department of Fisheries Technology, Bangladesh Agricultural University, Mymensingh-2202, Bangladesh"
}
],
"first_name": "Md. Mubarack",
"family_name": "Hossain",
"email": null,
"author_order": 2,
"ORCID": "http://orcid.org/0000-0001-7275-2429",
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 132
},
{
"id": 540,
"affiliation": [
{
"affiliation": "Department of Fisheries Technology, Bangladesh Agricultural University, Mymensingh-2202, Bangladesh"
}
],
"first_name": "Md. Naim",
"family_name": "Uddin",
"email": null,
"author_order": 3,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 132
},
{
"id": 541,
"affiliation": [
{
"affiliation": "Department of Fisheries Technology, Bangladesh Agricultural University, Mymensingh-2202, Bangladesh"
}
],
"first_name": "Md. Nurul",
"family_name": "Haider",
"email": "raselmnh@bau.edu.bd",
"author_order": 4,
"ORCID": "http://orcid.org/0000-0001-7656-7802",
"corresponding": true,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "Md. Nurul Haider, PhD; Department of Fisheries Technology, Bangladesh\r\nAgricultural University, Mymensingh-2202, Bangladesh e-mail: raselmnh@bau.edu.bd",
"article": 132
}
],
"views": 1530,
"downloads": 168,
"references": [
{
"id": 4328,
"serial_number": 1,
"pmc": null,
"reference": "DoF. Yearbook of Fish Stat Bangladesh, 2017-18 Fish resour surv syst (FRSS), Dep Fish Bangladesh Minist Fish. 2018;35:129.",
"DOI": null,
"article": 132
},
{
"id": 4329,
"serial_number": 2,
"pmc": null,
"reference": "Belton B, Karim M, Thilsted S, Murshed-E-Jahan K, Collis W, Phillips M. Aquaculture and fish consumption in Bangladesh Project Leader. 2011.",
"DOI": null,
"article": 132
},
{
"id": 4330,
"serial_number": 3,
"pmc": null,
"reference": "Bhuiyan ANMRK, Hossain MM, Uddin MN, Hossain MA, Hossain MI, Haider MN. Changes in viable bacterial counts and physicochemical parameters of water used during live transportation of Pangasius catfish (Pangasianodon hypophthalmus) in Bangladesh. J Adv Vet Anim Res. 2022;9:66–77.",
"DOI": null,
"article": 132
},
{
"id": 4331,
"serial_number": 4,
"pmc": null,
"reference": "Berka R. The transport of live fish. A review. EIFAC Technical Paper 1986; 48, 52 pp.",
"DOI": null,
"article": 132
},
{
"id": 4332,
"serial_number": 5,
"pmc": null,
"reference": "McFarland WNTK, Norris S. The control of pH by buffers in fish transport.” California Fish and Game. 1958;4: 291-310.",
"DOI": null,
"article": 132
},
{
"id": 4333,
"serial_number": 6,
"pmc": null,
"reference": "Hossain MM, Bhuiyan ANMRK, Hossain MA, Uddin MN, Hossain MI, Haider MN. Follow up of bacterial and physicochemical quality of water during live transportation of climbing perch (Anabas testudineus) in Bangladesh. J Adv Biotechnol Exp Ther. 2021;4:149–60.",
"DOI": null,
"article": 132
},
{
"id": 4334,
"serial_number": 7,
"pmc": null,
"reference": "Newaj-Fyzul A, Mutani A, Ramsubhag A, Adesiyun A. Prevalence of bacterial pathogens and their anti-microbial resistance in tilapia and their pond water in Trinidad. Zoonoses Public Health. 2008;55:206–13.",
"DOI": null,
"article": 132
},
{
"id": 4335,
"serial_number": 8,
"pmc": null,
"reference": "Wogu M, Maduakor C. Evaluation of microbial spoilage of some aquacultured fresh fish in benin city Nigeria. Ethiop J Environ Stud Manag. 2011;3.",
"DOI": null,
"article": 132
},
{
"id": 4336,
"serial_number": 9,
"pmc": null,
"reference": "Aziz H, Daphn A. Bacteriological studies of faecal and water samples from different sources with special reference to some Gram-negative bacteria. Benha Vet. Med. J.,2005:248-61.",
"DOI": null,
"article": 132
},
{
"id": 4337,
"serial_number": 10,
"pmc": null,
"reference": "FAO. Assessment and management of seafood safety and quality. FAO fisheries technical paper 2003:444.",
"DOI": null,
"article": 132
},
{
"id": 4338,
"serial_number": 11,
"pmc": null,
"reference": "Pao C, Molla B, Kleer J, Reine A. Hygienic control of fish processing plant. Wochenschr. 2008;121(4):89-93.",
"DOI": null,
"article": 132
},
{
"id": 4339,
"serial_number": 12,
"pmc": null,
"reference": "Traore O, Nyholm O, Siitonen A, Bonkoungou IJO, Traore AS, Barro N. Prevalenceand diversity of Salmonella enterica in water, fish and lettuce in Ouagadougou, Burkina Faso. BMC Microbiol 2015;15:1–7.",
"DOI": null,
"article": 132
},
{
"id": 4340,
"serial_number": 13,
"pmc": null,
"reference": "Ampofo JA, Clerk GC. Diversity of bacteria contaminants in tissues of fish cultured in organic waste-fertilized ponds: Health Implications~!2009-10-15~!2010-02-16~!2010-06-17~! Open Fish Sci J. 2010;3:142–6.",
"DOI": null,
"article": 132
},
{
"id": 4341,
"serial_number": 14,
"pmc": null,
"reference": "Esposto EM, Silva WCP, Reis CMF, Reis EMF, Ribeiro R V., Rodrigues DP. Enteropatogenos bacterianos em peixes criados em uma estacao de reciclagem de nutrientes e no ecossistema relacionado. Pesqui Veterinária Bras. 2007;27:144–8.",
"DOI": null,
"article": 132
},
{
"id": 4342,
"serial_number": 15,
"pmc": null,
"reference": "Li K, Petersen G, Barco L, Hvidtfeldt K, Liu L, Dalsgaard A. Salmonella weltevreden in integrated and non-integrated tilapia aquaculture systems in Guangdong, China. Food Microbiol. 2017;65:19–24.",
"DOI": null,
"article": 132
},
{
"id": 4343,
"serial_number": 16,
"pmc": null,
"reference": "Nwiyi P, Onyeabor A. Occurrence of Salmonella spp from fresh fish (Tilapia Nilotica Linn) using improved isolation methods. Online J Anim Feed Res. 2012;2:475–8.",
"DOI": null,
"article": 132
},
{
"id": 4344,
"serial_number": 17,
"pmc": null,
"reference": "Wong MH, Chen S. First detection of oqAB in Salmonella spp. isolated from food. Antimicrobial agents and chemotherapy. 2013;;57:658-60.",
"DOI": null,
"article": 132
},
{
"id": 4345,
"serial_number": 18,
"pmc": null,
"reference": "Awuor WS, Miruka OD, Eliud WN. Characterisation of Salmonella isolated from nile tilapia (Oreochromis niloticus) along lake victoria beaches in western Kenya. Inter J Biol Med Sciences. 2011;1:51-6.",
"DOI": null,
"article": 132
},
{
"id": 4346,
"serial_number": 19,
"pmc": null,
"reference": "Mhango M, Mpuchane SF, Mpuchane BA. Incidence of indicator organisms, opportunistic and pathogenic bacteria in fish. African Journal of Food, Agriculture, Nutrition and Development. 2010;10:10.",
"DOI": null,
"article": 132
},
{
"id": 4347,
"serial_number": 20,
"pmc": null,
"reference": "Wyatt LE, Nickelson R, Vanderzant C. Occurrence and control of Salmonella in freshwater catfish. Journal of Food Science. 1979;4:67-73.",
"DOI": null,
"article": 132
},
{
"id": 4348,
"serial_number": 21,
"pmc": null,
"reference": "Wyatt LE, Nickelson R, Vanderzant CA. Edwardsiella tarda in freshwater catfish and their environment. Applied and Environmental Microbiology. 1979;38:710-4.",
"DOI": null,
"article": 132
},
{
"id": 4349,
"serial_number": 22,
"pmc": null,
"reference": "Rompre A, Servais P, Baudart J, Roubin MR, Laurent P. Detection and enumeration of coliforms in drinking water: current methods and emerging approaches. Journal of microbiological methods. 2002;49:31-54.",
"DOI": null,
"article": 132
},
{
"id": 4350,
"serial_number": 23,
"pmc": null,
"reference": "Buras N, Duek L, Niv S, Hepher B, Sandbank E. Microbiological aspects of fish grown in treated wastewater. Water Res 1987;21:1–10.",
"DOI": null,
"article": 132
},
{
"id": 4351,
"serial_number": 24,
"pmc": null,
"reference": "Hejkal TW, Gerba CP, Henderson S, Freeze M. Bacteriological, virological and chemical evaluation of a wastewater-aquaculture system. Water Res 1983;17:1749–55.",
"DOI": null,
"article": 132
},
{
"id": 4352,
"serial_number": 25,
"pmc": null,
"reference": "Hejkal TW, Gerba CP, Henderson S, Freeze M. Bacteriological, virological and chemical evaluation of a wastewater-aquaculture system. Water Res 1983;17:1749–55.",
"DOI": null,
"article": 132
},
{
"id": 4353,
"serial_number": 26,
"pmc": null,
"reference": "Sarker S, Mahmud S, Sultana R, Biswas R, Sarkar PP, Munayem MA. Quality assessment of surface and drinking water of nakla paurosova, Sherpur, Bangladesh. Adv Microbiol 2019;09:703–27.",
"DOI": null,
"article": 132
},
{
"id": 4354,
"serial_number": 27,
"pmc": null,
"reference": "Ogunremi D, Davis S, Dupras AA, Marquez IG, Omidi K, Pope L. Evaluation of a multiplex pcr assay for the identification of Salmonella serovars enteritidis and typhimurium using retail and abattoir samples. J Food Prot 2017;80:295–301.",
"DOI": null,
"article": 132
},
{
"id": 4355,
"serial_number": 28,
"pmc": null,
"reference": "Hasan GMMA, Hossain MS, Parveen S, Juliana FM. Microbiological and chemical quality assessment of six fish species of Bangladesh during freeze storage. Int J Res Appl Sci Eng Technol 2016;4:572–8.",
"DOI": null,
"article": 132
},
{
"id": 4356,
"serial_number": 29,
"pmc": null,
"reference": "Haider MN, Faridullah M, Kamal M, Islam MN, Khan MN. A bacteriological assessment for Salmonella and Escherichia coli in some selected freshwater prawn (Macrobrachium rosenbergii) farms and depots. Journal of Marine Bioscience and Biotechnology. 2007;2:40-7.",
"DOI": null,
"article": 132
},
{
"id": 4357,
"serial_number": 30,
"pmc": null,
"reference": "Hossain M, Siddique M, Hossain F, Zinnah M, Hossain M, Alam M. Isolation, identification, toxin profile and antibiogram of Escherichia coli isolated from broilers and layers in Mymensingh district of Bangladesh. Bangladesh J Vet Med 1970;6:1–5.",
"DOI": null,
"article": 132
},
{
"id": 4358,
"serial_number": 31,
"pmc": null,
"reference": "Oh JY, Kang MS, Kim JM, An BK, Song EA, Kim JY. Characterization of Escherichia coli isolates from laying hens with colibacillosis on 2 commercial egg-producing farms in Korea. Poult Sci 2011;90:1948–54.",
"DOI": null,
"article": 132
},
{
"id": 4359,
"serial_number": 32,
"pmc": null,
"reference": "Yanestria SM, Rahmaniar RP, Wibisono FJ, Effendi MH. Detection of invA gene of Salmonella from milkfish (Chanos chanos) at Sidoarjo wet fish market, Indonesia, using polymerase chain reaction technique. Vet World 2019;12:170–5.",
"DOI": null,
"article": 132
},
{
"id": 4360,
"serial_number": 33,
"pmc": null,
"reference": "Trust TJ, Sparrow RA. The bacterial flora in the alimentary tract of freshwater salmonid fishes. Canadian Journal of Microbiology. 1974;20:1219-28.",
"DOI": null,
"article": 132
},
{
"id": 4361,
"serial_number": 34,
"pmc": null,
"reference": "[34] Mobin SM, Chowdhury MB, Islam MS, Uddin MN. Status of bacterial flora in the intestine of two freshwater fish. Bangladesh J. Life Sci. 2001;13:149-55.",
"DOI": null,
"article": 132
},
{
"id": 4362,
"serial_number": 35,
"pmc": null,
"reference": "Al-Harbi AH, Uddin MN. Seasonal variation in the intestinal bacterial flora of hybrid tilapia (Oreochromis niloticus x Oreochromis aureus) cultured in earthen ponds in Saudi Arabia. Aquaculture 2004;229:37–44.",
"DOI": null,
"article": 132
},
{
"id": 4363,
"serial_number": 36,
"pmc": null,
"reference": "Islam MS, Siddika A, Khan MNH, Goldar MM, Sadique MA, Kabir ANMH. Microbiological analysis of tube-well water in a rural area of Bangladesh. Appl Environ Microbiol 2001;67:3328–30.",
"DOI": null,
"article": 132
},
{
"id": 4364,
"serial_number": 37,
"pmc": null,
"reference": "John DE, Rose JB. Review of factors affecting microbial survival in groundwater. Environ Sci Technol 2005;39:7345–56.",
"DOI": null,
"article": 132
},
{
"id": 4365,
"serial_number": 38,
"pmc": null,
"reference": "Suthar S, Chhimpa V, Singh S. Bacterial contamination in drinking water: a case study in rural areas of northern Rajasthan, India. Environ Monit Assess 2009;159:43–50.",
"DOI": null,
"article": 132
}
]
},
{
"id": 131,
"slug": "178-1654949612-effect-of-gentamicin-and-doxycycline-on-expression-of-relb-and-rele-genes-in-klebsiella-pneumonia",
"featured": false,
"slider": false,
"issue": "Vol5 Issue3",
"type": "original_article",
"manuscript_id": "178-1654949612",
"recieved": "2022-06-11",
"revised": null,
"accepted": "2022-07-18",
"published": "2022-07-27",
"pdf_file": "https://jabet.bsmiab.org/media/pdf_file/2023/52/178-1654949612.pdf",
"title": "Effect of gentamicin and doxycycline on expression of relB and relE genes in Klebsiella pneumonia",
"abstract": "<p><em>Klebsiella pneumoniae</em> is responsible for a variety of disease in hospitalized patients. The goal of this study was to determine that <em>K. pneumoniae</em> isolates possessed toxin-antitoxin II genes such as <em>relE</em> and <em>relB</em>. Other than that, if there was a correlation between the expression of these two genes and antibiotic resistance in<em> K. pneumoniae. </em>Fifty-seven urine samples were collected from Baghdads’ hospitals; diagnosed and identified by phenotype and biochemical tests and confirmed with VITEK 2 compact system. Only fifteen isolates which were identified as <em>Klebsiella pneumoniae</em>. Antibiotic sensitivity was identified by using twelve antibiotics discs.<em> K. pneumoniae</em> showed 100% resistance to ceftriaxone, amoxicillin, ticarcillin, ticarcillin with clavulanic acid, ceftazidime, tetracycline, while other antibiotics showed less percent of resistant. Minimum inhibitory concentrations (MICs) of antibiotics detected by using macro tube dilution method to identify the antimicrobial activity for <em>K. pneumoniae</em>. The MIC of gentamicin and doxycycline antibiotics was 1024 Mg/ml, 512 Mg/ml, respectively. The <em>relB (</em>115 bp)<em>, </em>and <em>relE (</em>136 pb) genes were detected by polymerase chain reaction. Then gene expression of <em>relB and relE</em> was conducted by using (RT-qPCR) technique treated with sub-MIC concentration of (gentamicin and doxycycline) antibiotics. This study found only ten isolates harbored the two genes. The <em>relB </em>gene expression was increased, but at the same time <em>relE</em> gene expression was decreased compared to control <em>infB1</em> gene expression. This means the bacterial cell tolerance antibiotics sub-MIC concentrations by maintaining the number of bacteria under stress of antibiotics. Finally, these findings suggest the potential of <em>relB</em> to make<em> K. pneumoniae</em> resistant to antibiotics in their infections under antibiotic stress by the toxin-antitoxin II system.</p>",
"journal_reference": "J Adv Biotechnol Exp Ther. 2022; 5(3): 667-675.",
"academic_editor": "Md Jamal Uddin, PhD; ABEx Bio-Research Center, Dhaka-1230, Bangladesh",
"cite_info": "Sweedan EG , Shehab ZH, et al. Effect of gentamicin and doxycycline on expression of relB and relE genes in Klebsiella pneumonia. J Adv Biotechnol Exp Ther. 2022; 5(3): 667-675.",
"keywords": [
"Gentamicin",
"Gene expression",
"relE gene",
"Doxycycline",
"Klebseilla pneumoniae",
"relB gene"
],
"DOI": "10.5455/jabet.2022.d145",
"sections": [
{
"section_number": 1,
"section_title": "INTRODUCTION",
"body": "<p><em>Klebsiella pneumoniae </em>is a bacterial pathogen of major importance that causes a variety of disease manifestations in hospitalized patients [<a href=\"#r-1\">1</a>]. <em>K. pneumoniae </em>is rapidly generating multidrug resistance (MDR), posing a severe hazard to patients due to a higher mortality rate and lower therapeutic efficiency. <em>K. pneumoniae</em> can develop antibiotic resistance more rapidly than other bacteria due to the production of enzymes such as extended-spectrum ß-lactamases (ESBLs) and carbapenemase [<a href=\"#r-2\">2-4</a>]. Exposure to antibiotics is a major risk factor for developing antibiotic resistance in bacteria. The extensive and prolonged use of antibiotics is a crucial factor in the development of resistance in bacteria for diseases associated with healthcare. [<a href=\"#r-5\">5</a>].<br />\r\nA toxin-antitoxin (TA) system is a group of two or more tightly related genes of a protein that encode a poison and a cure. In the conventional physiology of bacteria, an antitoxin attaches to a toxin and neutralizes it, preventing the bacterium from killing itself. This system consists of two genes in an operon, one of which produces a stable toxin and the other of which produces a less stable antitoxin [<a href=\"#r-6\">6</a>].<br />\r\nThe frequency of toxins-antitoxins system especially type II in bacteria, as well as their involvement in their pathogenicity, biofilm formation, and bacteriophage resistance in these bacteria which have toxin-antitoxin II system. The biological activities of these systems have many functions, including roles in antibiotic resistance and bacterial persistence [<a href=\"#r-7\">7</a>].<br />\r\nThis study aimed to determine whether or not <em>K</em>. <em>pneumoniae</em> isolates possessed toxins-antitoxins II genes such as <em>relE </em>and <em>relB</em>. Other than that, if there was a correlation between the expression of these two genes and antibiotic resistance in <em>K. pneumoniae</em></p>"
},
{
"section_number": 2,
"section_title": "MATERIALS AND METHODS",
"body": "<p><strong>Bacterial isolation and identification</strong><br />\r\nFifteen <em>K. pneumoniae</em> were identified from fifty-seven samples of urine from patients by phenotyping and biochemical tests. The plates of MacConkey agar and blood agar were streaked with urine and then incubated at 37°C overnight. The bacteria showed pink colonies on MacConkey agar because of lactose fermentation. <em>K. pneumoniae</em> isolates showed the positive result for the Simmon citrate test indole test [<a href=\"#r-8\">8</a>]. Then confirmation of identification was done by VITEK 2 Compact system.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Antibiotics sensitivity test</strong><br />\r\nResistant of isolates was determined using the disc’s technique [<a href=\"#r-9\">9</a>] to twelve different antibiotics discs (Bioanalyse/Turkey): Ceftazidime(30µg), ceftriaxne (30µg), imipenem(10µg), amoxicillin (30µg), ciprofloxacin (10µg), ticarcillin (10µg), ticarcillin/clavulanic acid (75 µg /10µg), kanamycin(10µg), gentamicin (10µg) nitrofurantion (30µg), tetracycline(30µg), and doxycyciline (30µg). The isolated colony was cultured on nutrient broth overnight. Then it was cultured on Muller-Hinton agar after being diluted to 1.5×10<sup>8 </sup>(cell/ml). Discs of antibiotics were fixed on the cultured plates by sterile forceps. After that, the plate was incubated at 37°C overnight. Then the results were compared with CLSI data in 2019 [<a href=\"#r-10\">10</a>].</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Antibiotic minimum inhibitory concentrations (MIC)</strong><br />\r\nIsolates were tested for sensitivity by using the macro dilution broth assay, which was used to estimate the MIC of two antibiotics (doxycycline and gentamicin) for <em>K. pneumoniae</em> [<a href=\"#r-9\">9</a>]. After dilution to 1.5×10<sup>8</sup> (cell/ml), bacteria were inoculated on Mueller-Hinton broth using a sterile loop, and antibiotics with double serial concentrations were administered to the medium. Then the medium was incubated for 24 hours at 37°C. The results of MIC value were the first clear test tube after turbid tubes.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>PCR analysis</strong><br />\r\nThermal lysis and centrifugation method at 4°C for 30 seconds at 9,000 rpm were used to extract bacterial DNA. Nanodrop was used to assess the DNA content of the supernatant. Each step was completed in a final volume of 25 μl, by using 12.5 μl master mix, 1.5 μl primers and 100 ng DNA plus nuclease-free water. PCR amplification was used to identify <em>relE </em>and<em> relB</em> as previously described. The step of initial denaturation was done for 5 minutes at 95°C, while denaturation at 95°C for 30 sec. Then annealing was done at 62°C with the same time of previous step. After that the step of final extension was prepared at 72°C for 30 sec. Electrophoresis on 1.5% (w/v) agarose gel in tris acetate-EDTA (TBE) buffer resolved all PCR-amplified products. The samples were run at 100 V/Amp. for around 75 minutes. Using a conventional ultraviolet-light transilluminator, the specified products were detected following ethidium bromide staining [<a href=\"#r-11\">11</a>]. The study genes were amplified with specific primers (Macrogen, USA) for <em>relE: F</em>-GCACTAAAGGAATGGCGAAAG, R-GGAGCTTGTTTGCTTCAATCC; <em>relB:</em> F-AATGGGCGTAACTCCTTCTG, R-CACAAGTTCAGCATCTTCATCAC; and infB1: F-CTCGCTGCTGGACTATATTCG, R-CGCTTTCAGCTCAAGAACTTC.<br />\r\nThe same steps and conditions of PCR were used to investigate some toxin-antitoxin type II genes (<em>relE</em> and <em>relB</em>) and housekeeping genes. [<a href=\"#r-12\">12</a>].</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Gene expression of <em>relE</em> and <em>relB</em> genes by using Pfaffi method</strong><br />\r\nThe concentration of extracted RNA was measured using a Quantus Fluorometer to determine sample quality for downstream applications [<a href=\"#r-13\">13</a>]. 199 µl of diluted quantus flour dye was combined with 1 µl of RNA. RNA concentrations were measured after a 5-minute incubation period at room temperature in a dark environment. Macrogen Company provided the primers in lyophilized form. As a stock solution, lyophilized primers were dissolved in nuclease-free water to a final concentration of 100 pmol/μl. 10 μl of primer stock solution was mixed with 90 μl of nuclease-free water.<br />\r\nRNA was isolated from <em>K. pneumoniae</em> using trizol reagent (Promega, USA) as described in the protocol by the manufacturer. Gene expression of toxin-antitoxin genes was measured via the relative (RT-qPCR) technique. The same conditions and program as performed previously by one-step real-time PCR with use the same primer sets of genes. The <em>infB1 </em>was used as housekeeping primer in real-time PCR with the same program steps but the annealing temperature was 50°C.<br />\r\nThe gene expression was achieved with two positive isolates for these genes. And control isolate of <em>K. pneumoniae</em> without antibiotics treatment. Then analysis gene expression was calculated by using Pfaffi method for relative quantification as described previously [<a href=\"#r-14\">14, 15</a>].<br />\r\nFolding change = 2<sup>-ΔΔCT</sup><br />\r\nΔCT = CT <sub>(gene)</sub> – CT <sub>(House Keeping gene)</sub><br />\r\nΔΔCT = ΔCT <sub>(Treated) </sub>– ΔCT <sub>(Control)</sub></p>"
},
{
"section_number": 3,
"section_title": "RESULTS",
"body": "<p><strong>Effect of antibiotics on <em>K. pneumoniae</em></strong><br />\r\nFifteen<em> K. pneumoniae</em> were evaluated for antibiotic susceptibility to a variety of antibiotic types. <em>K. pneumoniae </em>isolates were 100 % resistant to ceftriaxone, amoxicillin, ticarcillin, ticarcillin with clavulanic acid, ceftazidime, and tetracycline, as shown in <a href=\"#figure1\">Figure 1</a>. While ciprofloxacin and nitrofurantoin showed intermediate resistance (75%). While all isolates were sensitive to imipinem (94.6%) and only 6.4% were resitant to it. But doxycycline and gentamicin showed 67.7% of resistant isolates, and 32.3% of intermediate and persistent were sensitive. But in amikacin 35% of the isolates were sensitive and 25% were intermediate, and 40% were resistant.</p>\r\n\r\n<div id=\"figure1\">\r\n<figure class=\"image\"><img alt=\"\" height=\"213\" src=\"/media/article_images/2023/40/25/178-1654949612-Figure1.jpg\" width=\"474\" />\r\n<figcaption><strong>Figure 1.</strong> Results of antibiotic sensitivity test percent for <em>K. pneumoniae</em> isolates 1: Amoxicillin, 2: Cefitrixone, 3: Ticarcillin, 4: Ticarcillin/Clavulanic acid, 5: Ceftazidim, 6: Tetracycline, 7: Ciprofloxcin, 8: Nitrofurantoin, 9: Gentamicin, 10: Doxycycline ,11: Imipenem, 12: Amikacin.</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Antibiotic minimum inhibitory concentrations</strong><br />\r\nThe macro tube dilution method was used to estimate the MIC for two antibiotics (doxycycline and gentamicin), when the first clear tube after serial of turbid tubes. The MICs value were 256 µg/ml for doxycycline and 1024 µg/ml for gentamicin as shows in <a href=\"#Table-1\">Table 1</a>.</p>\r\n\r\n<div id=\"Table-1\">\r\n<p><a href=\"https://jabet.bsmiab.org/table/178-1654949612-table1/\">Table-1</a><strong>Table 1.</strong> MICs value of antibiotics in <em>K. pneumoniae.</em></p>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Prevalence of <em>relE and relB</em> genes in the isolates</strong><br />\r\nThe prevalence of these genes in ten isolates is shown in <a href=\"#figure2\">Figure 2</a> and <a href=\"#figure3\">Figure 3</a>. The results showed that only ten isolates were positive with products 136bp (<em>relE</em>) and 115bp (<em>relB</em>)) when 67 % of isolates have two genes<em> relE </em>and<em> relB</em>.<br />\r\nThese results also confirmed the simultaneous presence of the toxin and antitoxin genes, as these genes are interrelated to neutralize the effect of the toxin on bacterial cells and prevent bacterial death.</p>\r\n\r\n<div id=\"figure2\">\r\n<figure class=\"image\"><img alt=\"\" height=\"195\" src=\"/media/article_images/2023/40/25/178-1654949612-Figure2.jpg\" width=\"442\" />\r\n<figcaption><strong>Figure 2. </strong>Results of the amplification of <em>relE</em> (136 bp.) on 1.5% agarose gel electrophoresis stained with Ethidium Bromide at 100 volts/Amp for 75 min, M: 100bp Marker.</figcaption>\r\n</figure>\r\n</div>\r\n\r\n<div id=\"figure3\">\r\n<figure class=\"image\"><img alt=\"\" height=\"180\" src=\"/media/article_images/2023/40/25/178-1654949612-Figure3.jpg\" width=\"442\" />\r\n<figcaption><strong>Figure 3. </strong>Results of the amplification of <em>relB</em> (115 bp.) on 1.5% agarose gel electrophoresis stained with Ethidium Bromide at 100 volts/Amp for 75 min, M: 100bp Marker.</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Effect of antibiotics on the expression of <em>relE, </em>and<em> relB</em> genes</strong><br />\r\nThe results showed that RNA concentrations ranged from 31, 29, and 25 ng/ml in an untreated isolate (K2) to 26, 5, and 11 ng/ml in isolates treated with antibiotics with sub-MICs (doxycycline and gentamicin). The expression of the gene was detected by using RT- qPCR using specific primer (housekeeping gene of <em>infB1</em>). The amplification accuracy of gene products was noticed by the value of cycle threshold (Ct), as shown in <a href=\"#figure3\">Figure 3</a>.<br />\r\nDue to the increasing incidence of novel resistant strains, it is critical to find inhibitors targeting <em>K. pneumoniae </em>to prevent infection by adopting alternate therapeutic strategies. The gene expression of <em>relE</em> and <em>relB</em> was studied after being treated with sub-MIC antibiotics. The data in <a href=\"#figure4\">Figures 4-6</a> showed that the <em>relB</em> gene was upregulated in treated isolates. The fold change in copy numbers between 8.4 and 15.2 was more than sixfold greater than the control. When reaching (0.7 to 0.9) copy numbers for <em>relE</em> gene in <a href=\"#figure7\">Figure 7</a>.</p>\r\n\r\n<div id=\"figure4\">\r\n<figure class=\"image\"><img alt=\"\" height=\"197\" src=\"/media/article_images/2023/40/25/178-1654949612-Figure4.jpg\" width=\"453\" />\r\n<figcaption><strong>Figure 4.</strong> Ct value of <em>infB1</em> in <em>K. pneumoniae</em> for treated and without treated with sub-MIC of antibiotics (doxycycline and gentamicin) as control.</figcaption>\r\n</figure>\r\n</div>\r\n\r\n<div id=\"figure5\">\r\n<figure class=\"image\"><img alt=\"\" height=\"199\" src=\"/media/article_images/2023/40/25/178-1654949612-Figure5.jpg\" width=\"441\" />\r\n<figcaption><strong>Figure 5.</strong> Ct value of <em>relB</em> in <em>K. pneumoniae</em> for treated and without treated with sub-MIC of antibiotics (doxycycline and gentamicin).</figcaption>\r\n</figure>\r\n</div>\r\n\r\n<div id=\"figure6\">\r\n<figure class=\"image\"><img alt=\"\" height=\"193\" src=\"/media/article_images/2023/40/25/178-1654949612-Figure6.jpg\" width=\"441\" />\r\n<figcaption><strong>Figure 6. </strong>Ct value of <em>relE</em> in <em>K. pneumoniae</em> for treated and without treated with sub-MIC of antibiotics (doxycycline and gentamicin).</figcaption>\r\n</figure>\r\n</div>\r\n\r\n<div id=\"figure7\">\r\n<figure class=\"image\"><img alt=\"\" height=\"213\" src=\"/media/article_images/2023/40/25/178-1654949612-Figure7.jpg\" width=\"441\" />\r\n<figcaption><strong>Figure 7. </strong>Fold changes in expression of <em>relB</em> and <em>relE</em> in <em>K. pneumoniae</em> isolate which treated with sub-MIC (doxycycline and gentamicin) antibiotics.</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>"
},
{
"section_number": 4,
"section_title": "DISCUSSION",
"body": "<p><em>K. pneumoniae</em> is considered one of the most important bacteria because it causes many lethal diseases in the world. This study found that fifteen <em>K. pneumoniae</em> isolates were antibiotic resistant, with each isolate displaying a varied percentage of resistance. In another study from 801 samples, 580 were urine whereas, 221 were pus and sputum. Samples were collected from patients between 6 months to 90 years of both sexes [<a href=\"#r-16\">16</a>]. Ten <em>K. pneumoniae </em>were isolated from urine and this outcome closely resembles the findings of the present study. They were 100% resistant to ampicillin; 70% to cefazoline, nitrofurantoin, and ofloxacin; 90% to cefotaxime and ceftriaxone; and 80% to ceftazidime. Imipenem followed by amikacin was the most effective antibiotic [<a href=\"#r-17\">17</a>].<br />\r\nThe genes <em>relE and relB</em> are type of toxin-antitoxin system type II and they are involved in antibiotics resistance [<a href=\"#r-18\">18</a>]. Although 10 isolates harbored two genes from fifteen isolates, many studies mentioned that<em> K. pneumoniae</em> had these genes. Two hundred and twelve putative type II TA loci were identified in 30 replicons of these <em>K. pneumoniae</em> strains [<a href=\"#r-19\">19</a>]. These results agree that the MIC concentration of the garlic extract can enhance the expression of the antitoxin gene since the bacteria preserve their numbers from death (programmed cell death) through decreasing gene expression of the toxin. This explains the possibility of using higher concentrations of MIC for used antibiotics [<a href=\"#r-17\">17</a>]. The expression of the antitoxin <em>relB</em> might reverse the toxin <em>relE</em>. These findings showed that while the creation of the <em>relE</em> toxin does not kill cells, it does cause cell immobility when cells are exposed to antibiotics. The expression of the <em>relB</em> antitoxin might reverse this state of rest. Both genes are expressed under normal or favorable conditions, allowing the toxin’s effects to be inhibited. In contrast, the antitoxins are swiftly destroyed by proteases under stress, leaving the more stable toxin to impact cell development, generally as ribonucleases [<a href=\"#r-18\">18</a>].<br />\r\nAnother research looked at gene expression in the presence of gentamicin sub-MIC and <em>relE1-relB1, hipA-hipB, doc-phd, </em>and<em> mazF-mazE </em>loci were upregulated. Whereas the <em>relE2-relB2</em> and <em>vapC-vapB</em> loci were downregulated. Since there is little information about the function of type II toxin-antitoxin systems in <em>K. pneumoniae</em> response to different stressors, the expression levels of TA system genes in <em>K. pneumoniae</em> were investigated under oxidative and antibiotic stress [<a href=\"#r-20\">20, 21</a>]. These findings showed that bacterial cells can withstand antibiotic exposure due to a long-term upregulation of relE. This mechanism is unknown, but bacteria may use it to survive antibiotics and other stresses.</p>"
},
{
"section_number": 5,
"section_title": "CONCLUSION",
"body": "<p>This study concluded that <em>relB</em> gene expression is downregulated in copy numbers compared to <em>relE</em> after being treated with antibiotics. This indicates the bacterial cell can tolerate antibiotics sub-MIC concentrations by maintaining their number under the stress of antibiotics. Thus, gentamicin and doxycycline antibiotics were used as a good treatment against <em>K. pneumoniae</em>. Further study is required on the toxin-antitoxin system and its function, as well as its relationship with antibiotic resistance in <em>K. pneumoniae.</em></p>"
},
{
"section_number": 6,
"section_title": "ACKNOWLEDGEMENT",
"body": "<p>The authors thank the technical support provided by Department of Biology / College of Science/University of Baghdad.</p>"
},
{
"section_number": 7,
"section_title": "AUTHORS CONTRIBUTION",
"body": "<p>MTF; was designed the experiments; performed the experiments; analysis and recorded data: SEG and ZHS was conceived and designed the experiments, analyzed the data, guided to draft the manuscripts, and improved accordingly.</p>"
},
{
"section_number": 8,
"section_title": "CONFLICTS OF INTEREST",
"body": "<p>There is no conflict of interest among the authors.</p>"
}
],
"figures": [
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/40/25/178-1654949612-Figure1.jpg",
"caption": "Figure 1. Results of antibiotic sensitivity test percent for K. pneumoniae isolates 1: Amoxicillin, 2: Cefitrixone, 3: Ticarcillin, 4: Ticarcillin/Clavulanic acid, 5: Ceftazidim, 6: Tetracycline, 7: Ciprofloxcin, 8: Nitrofurantoin, 9: Gentamicin, 10: Doxycycline ,11: Imipenem, 12: Amikacin.",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/40/25/178-1654949612-Figure2.jpg",
"caption": "Figure 2. Results of the amplification of relE (136 bp.) on 1.5% agarose gel electrophoresis stained with Ethidium Bromide at 100 volts/Amp for 75 min, M: 100bp Marker.",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/40/25/178-1654949612-Figure3.jpg",
"caption": "Figure 3. Results of the amplification of relB (115 bp.) on 1.5% agarose gel electrophoresis stained with Ethidium Bromide at 100 volts/Amp for 75 min, M: 100bp Marker.",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/40/25/178-1654949612-Figure4.jpg",
"caption": "Figure 4. Ct value of infB1 in K. pneumoniae for treated and without treated with sub-MIC of antibiotics (doxycycline and gentamicin) as control.",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/40/25/178-1654949612-Figure5.jpg",
"caption": "Figure 5. Ct value of relB in K. pneumoniae for treated and without treated with sub-MIC of antibiotics (doxycycline and gentamicin).",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/40/25/178-1654949612-Figure6.jpg",
"caption": "Figure 6. Ct value of relE in K. pneumoniae for treated and without treated with sub-MIC of antibiotics (doxycycline and gentamicin).",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/40/25/178-1654949612-Figure7.jpg",
"caption": "Figure 7. Fold changes in expression of relB and relE in K. pneumoniae isolate which treated with sub-MIC (doxycycline and gentamicin) antibiotics.",
"featured": false
}
],
"authors": [
{
"id": 535,
"affiliation": [
{
"affiliation": "Biology Department, College of Science, University of Baghdad, Baghdad, Iraq"
}
],
"first_name": "Enass Ghassan",
"family_name": "Sweedan",
"email": "enassghassan1@gmail.com",
"author_order": 1,
"ORCID": "http://orcid.org/0000-0001-7330-9939",
"corresponding": true,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "Enass Ghassan Sweedan, PhD; Biology Department, College of Science, University of Baghdad, Baghdad, Iraq, e-mail: enassghassan1@gmail.com",
"article": 131
},
{
"id": 536,
"affiliation": [
{
"affiliation": "Biology Department, College of Science for Women, University of Baghdad, Baghdad, Iraq"
}
],
"first_name": "Zina Hashem",
"family_name": "Shehab",
"email": null,
"author_order": 2,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 131
},
{
"id": 537,
"affiliation": [
{
"affiliation": "Biology Department, College of Science, University of Baghdad, Baghdad, Iraq"
}
],
"first_name": "May Talib",
"family_name": "Flayyih",
"email": null,
"author_order": 3,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 131
}
],
"views": 1085,
"downloads": 178,
"references": [
{
"id": 4307,
"serial_number": 1,
"pmc": null,
"reference": "Fodah RA, Scott JB, Tam H-H, Yan P, Pfeffer TL, et al. (2014) Correlation of Klebsiella pneumoniae Comparative Genetic Analyses with Virulence Profiles in a Murine Respiratory Disease Model. PLoS ONE 9(9): e107394. doi: 10.1371/journal.pone.0107394.",
"DOI": null,
"article": 131
},
{
"id": 4308,
"serial_number": 2,
"pmc": null,
"reference": "Podschun R, Ullmann U. Klebsiella spp. as nosocomial pathogens: epidemiology, taxonomy, typing methods, and pathogenicity factors. Clin Microbiol Rev. 1998 Oct;11(4):589-603. doi: 10.1128/CMR.11.4.589.",
"DOI": null,
"article": 131
},
{
"id": 4309,
"serial_number": 3,
"pmc": null,
"reference": "Bengoechea JA, Sa Pessoa J. Klebsiella pneumoniae infection biology: living to counteract host defences. FEMS Microbiol Rev. 2019 Mar 1;43(2):123-144. doi: 10.1093/femsre/fuy043.",
"DOI": null,
"article": 131
},
{
"id": 4310,
"serial_number": 4,
"pmc": null,
"reference": "Munita JM, and Arias CA. Mechanisms of antibiotic resistance. Microbiol Spectr. 2016;4(2). https://doi.org/10.1128/microbiolspec.VMBF-0016-2015.",
"DOI": null,
"article": 131
},
{
"id": 4311,
"serial_number": 5,
"pmc": null,
"reference": "Prestinaci F, Pezzotti P, Pantosti A. Antimicrobial resistance: a global multifaceted phenomenon. Pathog Glob Health. 2015;109(7):309-18. doi: 10.1179/2047773215Y.0000000030. Epub 2015 Sep 7. PMID: 26343252; PMCID: PMC4768623.",
"DOI": null,
"article": 131
},
{
"id": 4312,
"serial_number": 6,
"pmc": null,
"reference": "Van Melderen L, Bernard P, Couturier M. Lon-dependent proteolysis of CcdA is the key control for activation of CcdB in plasmid-free segregant bacteria. Mol Microbiol. 1994 Mar;11(6):1151-7. doi: 10.1111/j.1365-2958.1994.tb00391.x.",
"DOI": null,
"article": 131
},
{
"id": 4313,
"serial_number": 7,
"pmc": null,
"reference": "Kamruzzaman M, Wu AY, Iredell JR. Biological Functions of Type II Toxin-Antitoxin Systems in Bacteria. Microorganisms. 2021 Jun 11;9(6):1276. doi: 10.3390/microorganisms9061276.",
"DOI": null,
"article": 131
},
{
"id": 4314,
"serial_number": 8,
"pmc": null,
"reference": "Collee JG, Miles RS, Watt B. Test for the identification of bacteria. In: Collee. J. G.; Fraser, A. G.; Marmion, B. P. and Simmons, A. (Eds.). Practical Medical Microbiology. 14P th P Edition. Churchill Livingstone, New York. pp. 131-146. 1996.",
"DOI": null,
"article": 131
},
{
"id": 4315,
"serial_number": 9,
"pmc": null,
"reference": "Kirby WMM, Bauer AW, Sherhis JC and Turck M. Antibiotic susceptibility testing by a standardized single disc method. Amer. J. Clin. Path. 1966; 45:493-496.",
"DOI": null,
"article": 131
},
{
"id": 4316,
"serial_number": 10,
"pmc": null,
"reference": "Clinical and Laboratory Standards Institute (CLSI). Performance standards for antimicrobial susceptibility testing. 29th ed. Clinical and Laboratory Standards Institute (CLSI). Supplement M100; 2019.",
"DOI": null,
"article": 131
},
{
"id": 4317,
"serial_number": 11,
"pmc": null,
"reference": "Ellington MJ, Kistler J, Livermore DM, Woodford N. Multiplex PCR for rapid detection of genes encoding acquired metallo-beta-lactamases. J Antimicrob Chemother. 2007 Feb;59(2):321-2. doi: 10.1093/jac/dkl481. Epub 2006 Dec 21. PMID: 17185300.",
"DOI": null,
"article": 131
},
{
"id": 4318,
"serial_number": 12,
"pmc": null,
"reference": "Narimisa N, Amraei F, Kalani BS, Mohammadzadeh R, Jazi FM. Effects of sub-inhibitory concentrations of antibiotics and oxidative stress on the expression of type II toxin-antitoxin system genes in Klebsiella pneumoniae. J Glob Antimicrob Resist. 2020 June; 21:51-56. doi: 10.1016/j.jgar.2019.09.005. Epub 2019 Sep 11. PMID: 31520807.",
"DOI": null,
"article": 131
},
{
"id": 4319,
"serial_number": 13,
"pmc": null,
"reference": "chmittgen, T., Livak, K. Analyzing real-time PCR data by the comparative CT method. Nat Protoc 3 ;2008 :1101–1108. https://doi.org/10.1038/nprot.2008.73",
"DOI": null,
"article": 131
},
{
"id": 4320,
"serial_number": 14,
"pmc": null,
"reference": "Shehab ZH, AL-Rubaii BAL. Effect of D-Mannose on Gene Expression of Neuraminidase Produced from Different Clinical Isolates of Pseudomonas aeruginosa. Baghdad Science Journal.2019; 16(2):291-298. DOI: http://dx.doi.org/10.21123/bsj.2019.16.2.0291",
"DOI": null,
"article": 131
},
{
"id": 4321,
"serial_number": 15,
"pmc": null,
"reference": "Luma Saeed Mohammed, Enass Ghassan Sweedan, May Talib Flayyih. Effects of Alcoholic Extracts of Cinnamomum zeylanicum and Origanum Majorana on Expression of Hly Gene in Escherichia coli. Indian Journal of Forensic Medicine & Toxicology.2020; 14(3):937-942.",
"DOI": null,
"article": 131
},
{
"id": 4322,
"serial_number": 16,
"pmc": null,
"reference": "Sweedan Enass Ghassan. Estimate Antimicrobial activity and Antibiofilm formation of bark (Cinnamomum zeylanicum) on Klebsiella pneumoniae from urinary tract infections. Iraqi Journal of Science. 2018; 59(3C): 1560-1566. DOI:10.24996/ijs.2018.59.3C.3",
"DOI": null,
"article": 131
},
{
"id": 4323,
"serial_number": 17,
"pmc": null,
"reference": "Fernández-García L, Blasco L, Lopez M, Bou G, García-Contreras R, Wood T, Tomas M. Toxin-Antitoxin Systems in Clinical Pathogens. Toxins (Basel). 2016 Jul 20;8(7):227. doi: 10.3390/toxins8070227. PMID: 27447671; PMCID: PMC4963858.",
"DOI": null,
"article": 131
},
{
"id": 4324,
"serial_number": 18,
"pmc": null,
"reference": "Zina Hashem Shehab, Enass Ghassan Sweedan, May Talib Flayyih. Evaluation the effect of Allium sativum (garlic) oil on the expression of mazE and mazF genes in Escherichia coli clinical isolates. Biochem. Cell. Arch. 2021;21(1):721-726.",
"DOI": null,
"article": 131
},
{
"id": 4325,
"serial_number": 19,
"pmc": null,
"reference": "Wei YQ, Bi DX, Wei DQ, Ou HY. Prediction of Type II Toxin-Antitoxin Loci in Klebsiella pneumoniae Genome Sequences. Interdiscip Sci. 2016 Jun;8(2):143-149. doi: 10.1007/s12539-015-0135-6. Epub 2015 Dec 10. PMID: 26662948.",
"DOI": null,
"article": 131
},
{
"id": 4326,
"serial_number": 20,
"pmc": null,
"reference": "Lieven Buts, Jurij Lah, Minh-Hoa Dao-Thi, Lode Wyns, and Remy Loris. Toxin–antitoxin modules as bacterial metabolic stress managers. TRENDS in Biochemical Sciences.2005;30 (12):672-679.",
"DOI": null,
"article": 131
},
{
"id": 4327,
"serial_number": 21,
"pmc": null,
"reference": "Negar Narimisa,Fatemeh Amraei, Behrooz Sadeghi Kalani. Biofilm establishment, biofilm persister cell formation, and relative gene expression analysis of type II toxin-antitoxin system in Klebsiella pneumoniae. Gene Reports. 2020; 21(2):100846. DOI: 10.1016/j.genrep.2020.100846.",
"DOI": null,
"article": 131
}
]
},
{
"id": 125,
"slug": "178-1655357396-effect-of-growth-factors-present-in-serum-on-insulin-resistance-and-endothelial-dysfunction-in-eahy926-cells",
"featured": false,
"slider": false,
"issue": "Vol5 Issue3",
"type": "original_article",
"manuscript_id": "178-1655357396",
"recieved": "2022-06-16",
"revised": null,
"accepted": "2022-07-18",
"published": "2022-07-26",
"pdf_file": "https://jabet.bsmiab.org/media/pdf_file/2023/12/178-1655357396.pdf",
"title": "Effect of growth factors present in serum on insulin resistance and endothelial dysfunction in EA.hy926 cells",
"abstract": "<p>Insulin resistance is a pathophysiological function of Type II diabetes mellitus which can be comprehended by quantifying the parameters critical to the insulin signaling pathway. Serum has a profound role in evaluating cellular growth and metabolism <em>in vitro</em>. We hypothesize that the growth factors present in serum such as IGF, EGF, and FGF have an effect on the regulatory components of the insulin signaling pathway leading to insulin resistance. This study focuses on the metabolic effect of Fetal Bovine Serum (FBS) in endothelial (EA.hy926) cells. A dose and time-dependent treatment of FBS on the chosen cells was followed by assessing cell viability and glucose uptake capacity using MTT and 2-NBDG assays respectively. Spectrophotometric analysis of nitric oxide (NO) and lactate dehydrogenase (LDH) determined vascular homeostasis and no cytotoxic effects of the serum, respectively, in endothelial cells. These findings indicate that FBS at higher levels could possibly lead to loss of NO activity which in turn could impair endothelium-mediated dilation. The inhibition of the enzymatic activity of eNOS, which in response to the stress may activate the release of LDH in endothelial cells displaying cytotoxicity. In conclusion, our findings indicate that a specific concentration of serum exposure enhances insulin signaling and endothelium cell regulation by modulating glucose uptake and NO production.</p>",
"journal_reference": "J Adv Biotechnol Exp Ther. 2022; 5(3): 623-633.",
"academic_editor": "Md Jamal Uddin, PhD; ABEx Bio-Research Center, Dhaka-1230, Bangladesh",
"cite_info": "Maganti H, Thandapani P, et al. Effect of growth factors present in serum on insulin resistance and endothelial dysfunction in EA.hy926 cells. J Adv Biotechnol Exp Ther. 2022; 5(3): 623-633.",
"keywords": [
"endothelial dysfunction",
"Insulin resistance",
"Growth factors",
"Fetal bovine serum",
"Dose-dependent"
],
"DOI": "doi.org/10.5455/jabet.2022.d141",
"sections": [
{
"section_number": 1,
"section_title": "INTRODUCTION",
"body": "<p>Type Ⅱ diabetes mellitus (T2DM) is a syndrome caused by a combination of metabolic impairments such as insulin receptor (IR) action, insulin receptor substrate (IRS), and phosphatidylinositol 3 kinase (PI3K) signaling. These aspects individually or in amalgamation with the others are devoted to the deterioration of vascular function and possibly lead to diabetic complications such as loss of endothelial cell function. A broad group of secreted polypeptides that are relatively small and stable are known as the growth factors [<a href=\"#r-1\">1</a>]. They play a significant role in cell growth, proliferation, differentiation, and cell signaling. Through the activation of specific transmembrane receptors in cells, relaying of the stimuli of the growth factor into the cell occurs [<a href=\"#r-2\">2</a>]. Serum is a powerhouse of several growth factors utilized in cell proliferation, differentiation, and growth. Fetal bovine serum (FBS) is inarguably used as an essential growth supplement in animal cell culture media and consists of several growth factors – EGF, FGF, VEGF, and IGF [<a href=\"#r-3\">3</a>]. We aim to investigate the effect of these metabolic factors on insulin signaling cascades. Even though prior studies have established the signaling effect of growth factors in causing insulin resistance, the metabolic aspect of the factors present in the serum along with the media is yet to be explored. Insulin is one among the factors which upon stimulation causes the phosphorylation of the insulin receptor substrate thereby leading to the activation of various downstream signaling aspects [<a href=\"#r-4\">4</a>]. Insulin also is responsible for the dilation of blood vessels and improves the overall flow of blood throughout the body. These blood vessels consist of a group of cells, including endothelial cells (EC) and vascular smooth muscle cells (VSMC) which coordinate and regulate vaso homeostasis [<a href=\"#r-5\">5</a>]. Endothelial cells have a vasomotor function in blood vessels, so the coordination of EC and VSMC maintains and regulates vaso homeostasis. For this process, Endothelial nitric oxide synthase (eNOS) acts as a marker of endothelial cell functions and produces NO, which undergoes paracrine signaling with associated cells in blood vessels. eNOS is the third isoform of the nitric oxide synthase gene (NOS3) and is one of the two constitutive isoforms of the gene [<a href=\"#r-6\">6</a>]. Nitric Oxide (NO) (a vasodilator in the endothelium) development is induced by Insulin which brings about a boost in blood circulation and decreased glucose uptake [<a href=\"#r-7\">7</a>]. At a cellular level, the PI3K-dependent insulin signaling pathway regulates endothelial-NO production by a potential enzymatic activity of endothelial Nitric oxide synthase (eNOS) [<a href=\"#r-8\">8</a>]. L-arginine is converted to NO by the eNOS expression. Additionally, it results in the regulation of vasoconstrictor endothelin-1 (ET-1) by the MAPK-dependent insulin signaling pathway [<a href=\"#r-9\">9</a>].<br />\r\nThis study focuses on serum (FBS), which in a concentration-dependent manner may ameliorate insulin resistance and develop a compensatory mechanism to enhance the signaling aspects of insulin-stimulated events such as glucose uptake, vaso homeostasis and metabolism in EA.hy926 endothelial cells. The specific aims were achieved by performing a time course study following cell viability and cytotoxicity assessment. Also, the metabolic effect on cells was determined by observing the glucose uptake capacity and levels of nitric oxide.</p>"
},
{
"section_number": 2,
"section_title": "MATERIALS AND METHODS",
"body": "<p><strong>Reagents and chemicals</strong><br />\r\nInsulin, Fetal Bovine Serum (FBS), and Dulbecco’s Modified Eagle Medium (DMEM), LDH assay kit were procured from HiMedia, India. Other reagents such as MTT (Thiazolyl Blue Tetrazolium Bromide), cell culture tested from HiMedia, <em>N</em><em><sup>ω</sup></em><em>-nitro-l-arginine methyl ester </em>(L-NAME) <u>></u> 98% TLC, powder from Sigma-Aldrich, (N-(1-Naphthyl) ethylenediamine dihydrochloride, Hi-AR/ACS) (NED) from HiMedia, and Sulfanilamide, Hi-AR<sup>TM</sup> <u>></u> 99% from HiMedia were purchased.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Cell culture of EA.hy926 cells</strong><br />\r\nEA.hy926 cells (passage no. 2) were obtained from HiMedia, India and were maintained at 37℃ in a CO<sub>2</sub> incubator. The cells were grown in DMEM with 10% Fetal Bovine Serum. The EA.hy926 were procured at passage number 2 and it takes 2 -3 days to reach 75% confluency. EA.hy926 cells, after reaching 75% confluence in three days were used for the entire study.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Cell viability test</strong><br />\r\nFollowing a 2-hour period of serum starvation, the viability of the cells with respect to varying serum concentrations (5%,10%,15% for 5min, 15min and 30min each) was analyzed by MTT [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide] assay. Triton X-100 was used as the positive control and DMSO was used as the solubilization buffer to dissolve the purple colour formazan product. Percentage cell viability was calculated after measuring the absorbance at 570nm [<a href=\"#r-10\">10</a>].</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Nitric oxide estimation</strong><br />\r\nThe nitric oxide test for endothelial cells was performed with the help of the Griess assay. The supernatant from the cell viability test was stored and used. A red-pink color develops after the addition of the NED (N-(1-Naphthyl) ethylenediamine) and sulfanilamide solutions. The intensity of this colour was measured at an absorbance of 540nm. The percentage of nitric oxide was calculated using the OD and slope values [<a href=\"#r-11\">11</a>].</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Cytotoxicity estimation by the release of lactate dehydrogenase</strong><br />\r\nThe cell cytotoxicity of EA.hy926 cells were estimated with the help of LDH assay with varying serum (FBS) concentrations (5%, 10%, 15%) and varying time intervals (5min, 15min, 30min). Triton X-100 (0.01%) served as the positive control and the control (untreated cells) served as negative control. The initial solution-A and stop solution that was added in the dark to the supernatant were used from the LDH assay kit, after which the absorbance was measured at 580nm. Percentage cytotoxicity was then calculated from the OD values obtained [<a href=\"#r-12\">12</a>].</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Measurement of 2-NBDG glucose uptake</strong><br />\r\nThe glucose uptake for endothelial cells was measured using a 2-NBDG (D-Glucose, 2-deoxy-2-((7-nitro-2,1,3-benzoxadiazol-4-yl) amino)-glucose) assay, with 2-NBDG being a glucose analog. Insulin was used as a positive control. The glucose uptake was measured with respect to the varying serum concentrations (5%, 10%, 15%) with varying time intervals. Excitation wavelengths of 485 or 355nm and emission wavelengths of 538 or 460nm were used to test fluorescence intensity [<a href=\"#r-13\">13</a>].</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Statistical analysis</strong><br />\r\nMean ± SEM were represented. The statistical significance was evaluated using one-way ANOVA in GraphPad Prism 8.0.2 to determine differences among groups. The criterion for statistical significance was *p < 0.05 and #p < 0.001.</p>"
},
{
"section_number": 3,
"section_title": "RESULTS",
"body": "<p><strong>Effect of serum treatment on cell viability</strong><br />\r\nAs described above, our initial aim was to determine the effect of exposure time and concentration of the FBS on the cells. The viability assay performed using MTT established that 15% FBS displayed maximum live cell count at each time point – 5 min, 15 mins and 30 mins. However, as the difference between cell viability among the concentrations and time points remained insignificant (p>0.05), it was concluded that the serum did not have a time and dose-dependent effect on the viability of EA. hy926 cells in comparison to the positive control, Triton-X 100 (<a href=\"#figure1\">Figure 1</a>).</p>\r\n\r\n<div id=\"figure1\">\r\n<figure class=\"image\"><img alt=\"\" height=\"353\" src=\"/media/article_images/2023/39/25/178-1655357396-Figure1.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 1.</strong> Assessment of cell viability by MTT in endothelial cells after treatment with fetal bovine serum in a time (5 min, 10min, 15min) and dose (5%, 10%, 15%) dependent manner. Triton X-100 (0.01%) is a positive control and DMSO serves as solvent control. Data represents the mean ± SEM of triplicates (N=3). #p < 0.001 as compared to control.</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Effect of serum treatment on nitric oxide levels</strong><br />\r\nThe endothelial cells following a dose and time-dependent FBS treatment were subjected to eNOS inhibitor L-NAME to compare the production of nitric oxide by untreated cells. The graph in <a href=\"#figure2\">Figure 2</a> shows that the production of NO is significantly reduced in the cells treated with L-NAME whereas the levels in basal cells remained unchanged.</p>\r\n\r\n<div id=\"figure2\">\r\n<figure class=\"image\"><img alt=\"\" height=\"397\" src=\"/media/article_images/2023/39/25/178-1655357396-Figure2.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 2. </strong>Nitric Oxide estimation in endothelial cells after treatment with fetal bovine serum in a time (5 min, 10min, 15min) and dose (5%, 10%, 15%) dependent manner, by using NED and sulphanilamide reagents. L-NAME is a positive control. Data represents the mean ± SEM of triplicates (N=3). *p < 0.05 and #p < 0.001 as compared to control.</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Effect of serum treatment on cell cytotoxicity</strong><br />\r\nThe endothelial cells were treated with FBS in a dose and time-dependent manner. The amount of LDH produced is increased when cells are exposed to Triton X-100 which is used as a positive control. It can be observed from the graph in <a href=\"#figure3\">Figure 3</a> that all the values are similar to the control and hence it can be concluded that FBS does not have any significant cytotoxic effect on the endothelial cells.</p>\r\n\r\n<div id=\"figure3\">\r\n<figure class=\"image\"><img alt=\"\" height=\"389\" src=\"/media/article_images/2023/39/25/178-1655357396-Figure3.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 3. </strong>LDH estimation in endothelial cells to observe cell cytotoxicity after treatment with fetal bovine serum in a time (5 min, 10min, 15min) and dose (5%, 10%, 15%) dependent manner. Triton X-100 (0.01%) is the positive control. Data represents the mean ± SEM of triplicates (N=3). #p < 0.001 as compared to control.</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Effect of serum treatment on 2-NBDG glucose uptake</strong><br />\r\nThe endothelial cells were exposed to FBS in a time and dose-dependent method. To ascertain the intracellular glucose accretion, a fluorescent glucose analogue (2-NBDG) was utilized to observe the uptake in individual cells [<a href=\"#r-12\">12</a>]. In <a href=\"#figure4\">Figure 4</a>, in the case of positive control due to insulin, we observe higher uptake of glucose in the cells.<br />\r\nThe percentage of glucose uptake by endothelial cells gradually increases, which were treated with serum, suggesting that as the concentration and incubation time increases a significant increase is observed in the uptake of glucose by the cells.</p>\r\n\r\n<div id=\"figure4\">\r\n<figure class=\"image\"><img alt=\"\" height=\"307\" src=\"/media/article_images/2023/39/25/178-1655357396-Figure4.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 4. </strong>Estimation of percentage of glucose uptake by 2-NBDG which is a fluorescent analogue of glucose, in endothelial cells after treatment with fetal bovine serum in a time (5 min, 10min, 15min) and dose (5%, 10%, 15%) dependent manner. Insulin (100nM) is a positive control and DMSO (0.01%) serves as solvent control. Data represents the mean ± SEM of triplicates (N=3). *p < 0.05 as compared to control.</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>"
},
{
"section_number": 4,
"section_title": "DISCUSSION",
"body": "<p>In this study we provide conclusive biological evidence that treatment of endothelial cells with serum at three serial concentrations trigger GLUT-4 activation followed by glucose uptake and eNOS enzymatic activity for endothelial cells function [<a href=\"#r-14\">14</a>]. We demonstrated that serum does not impact cell viability, whereas increased concentrations of serum was found to be non-cytotoxic to EA. hy926 cells.<br />\r\nWith insulin-induced eNOS phosphorylation and the reduction of IRS-2 expression, impaired insulin signaling in the endothelial cells reduces insulin-induced glucose uptake by the skeletal muscle via, at least in part, decreased capillary recruitment and decreased interstitial insulin concentrations in the skeletal muscle. As it happens, insulin signaling defect induced by IRS-2 deletion from the endothelial cells simultaneously impedes insulin-induced glucose uptake by the skeletal muscle, escalation in concentration of interstitial insulin and debilitation of the insulin-induced capillary recruitment. This insight into the mechanism also sheds light on the physiological roles of IRS-2 in the endothelial cells. Normal glucose homeostasis crucially seems to accommodate expression of a sufficient amount of IRS-2 in the endothelial cells. While there is surplus in IRS-2 expression during the fasting state, induction of sufficient glucose uptake by the skeletal muscle takes place and after feeding the raised glucose levels return to within the normal range. However, during the fasting period when endothelial cell expression of IRS-2 is decreased in the presence of hyperinsulinemia with insulin resistance, impairment in insulin signaling occurs, and the elevated levels of glucose after feeding fail to reduce efficaciously. Our results demonstrates that a fluorometric assay that is used to determine the glucose taken up by the cells in-turn aids in analyzing the cell metabolism as well as signaling characteristics. Thereby it is a mechanistic study of obesity and diabetes [<a href=\"#r-15\">15</a>]. The percentage of glucose absorption by serum-treated endothelial cells, suggesting that as the concentration and incubation time increases a significant increase is observed in the uptake of glucose by the cells. This can be explained by the improved expression of the glucose transporters such as GLUT-4. Insulin stimulates the mobilization of vesicles containing GLUT-4 glucose transporters to the plasma membrane, resulting in glucose transfer into the cells. In case of positive control due to hyperinsulinemia, caused by insulin dysfunction, higher uptake of glucose is observed in the cells. As a result, we may infer that a higher serum concentration aids in the reduction of insulin resistance by improving glucose uptake activity.<br />\r\nGenerally, both vasorelaxation and vasoconstriction are induced by insulin. Mediation of insulin-induced vasorelaxation is carried out by the IRS-PI3K-AKT pathway elevating endothelial NO production, whereas Shc/SOS/Ras-MAPK pathway inducing ET-1 expression mediates insulin-induced vasoconstriction [<a href=\"#r-16\">16</a>]. One of the principal features of the pathophysiology of diabetes is oxidative stress. Glycolysis metabolite methylglyoxal (MG) was formerly observed to affect redox balance by stimulating ROS production and modulating the expression and function of cytoprotective molecules such as glutathione [<a href=\"#r-17\">17</a>], superoxide dismutase [<a href=\"#r-18\">18</a>], glyoxalase [<a href=\"#r-19\">19</a>] and H<sub>2</sub>S [<a href=\"#r-20\">20</a>]. Increased NADPH oxidase-derived O<sub>2</sub><sup>•− </sup>reacts with eNOS-derived NO in order to create ONOO− that in turn oxidizes BH4 to BH2. In variety of conditions such as hyperphenylalaninemia, pathogenesis implies decrease in BH4 [<a href=\"#r-21\">21</a>]. Improvement was observed in diabetes [<a href=\"#r-22\">22</a>], ischemia reperfusion injury [<a href=\"#r-23\">23</a>], hypertension [<a href=\"#r-24\">24</a>], Alzheimer disease and Parkinson’s disease [<a href=\"#r-25\">25</a>], and in many of these pathological conditions due to exogenous BH4 [<a href=\"#r-21\">21,23</a>] or sepiapterin [<a href=\"#r-26\">26,27, 28, 29</a>] supplementation. BH4 is key in eNOS dimers maintenance and furthermore is functionally related to S-glutathionylation, a powerful eNOS uncoupling regulator [<a href=\"#r-29\">29</a>]. Our study demonstrates that Nitric oxide is considered one of the main regulators of energy metabolism and body composition and hence, measuring the amount of nitric oxide will help determine the condition and function of the cells [<a href=\"#r-30\">30</a>]. Although no significant conclusions can be derived based on the results of the cells treated with serum, a possible hypothesis could be made which explains that increased concentrations of serum might decrease the development of Nitric Oxide. Studies have shown that a drop in NO production from L-arginine in endothelial cells is a major factor for insulin resistance in diabetic patients [<a href=\"#r-31\">31,32</a>]. This can be explained by the idea that the cocktail of growth factors contained in the serum such as EGF, FGF and VEGF may have an adverse effect on the eNOS enzymatic activity leading to the depletion in NO production when compared to control. Endothelial dysfunction resulting in a decreased level of NO activity can not only guide vasoconstriction but can also impair endogenous negative feedback loop resulting in the inhibition of expression of certain pro-inflammatory cytokines [<a href=\"#r-33\">33</a>]. This decreased bioavailability of NO could be responsible for endothelial dysfunction which can be confirmed by performing further gene expression studies of certain specific targets [<a href=\"#r-8\">8</a>].<br />\r\nFBS serves as an exogenous source of LDH, which may have a significant consequence for cytotoxicity analysis [<a href=\"#r-30\">30</a>]. LDH, whose major role is to guide cellular respiration, is also associated with endothelial-derived soluble adhesion molecules and recognizes markers of endothelial activation that are commonly suppressed by NO. LDH is let into the bloodstream or into other bodily fluids when damage to tissues, organs, or the cells occurs. Similarly, when the cells are exposed to serum at three different concentrations for three different time intervals, in any case of the damage to the cells, there occurs a possibility for LDH to be released. Over expression of LDH may often interfere with normal glucose metabolism and insulin secretion by beta cells, and thus may be directly responsible for insulin secretory defects in some diabetes mellitus II cases [<a href=\"#r-34\">34</a>]. Testing the amount of LDH in the supernatant will establish the extent of cytotoxic effect of the serum on cells [<a href=\"#r-35\">35</a>]. From the results obtained, we can conclude that the amount of LDH produced is maximum when the endothelial cells are exposed to Triton X-100 which is used as a positive control. Among all the cells treated at different concentrations for different time periods, the ones treated for 30 minutes with a serum concentration of 15% exhibited higher LDH levels. However, these levels are not sufficient to be considered cytotoxic to the cells and hence, do not show proof of the serum possibly causing an insulin resistant condition due its high concentrations.<br />\r\nA cross talk study provides evidence stating that the insulin-mediated AKT and eNOS activations are induced optimally in the endothelial cells after feeding, resulting in insulin-induced capillary recruitment, interstitial insulin concentration increase, as well as elevation of skeletal muscle glucose. Due to inadequacy in the insulin-mediated AKT and eNOS activations in the endothelial cells of obese subjects after feeding, the insulin-induced capillary recruitment, interstitial insulin concentration increases and raise of skeletal muscle glucose uptake are damaged. With this information our study emphasized that serum does not have an impact on GLUT-4 translocation followed by glucose uptake, as understood at the cellular level of eNOS function on endothelial cells. However, the importance of insulin in balancing metabolic activity and endothelial cells functions and its crosstalk is remains unexplored at molecular level.</p>"
},
{
"section_number": 5,
"section_title": "CONCLUSION",
"body": "<p>This study focuses on observing if serum (FBS) might ameliorate insulin resistance and develop a compensatory mechanism to enhance the signaling aspects of insulin-stimulated events such as glucose uptake, vaso homeostasis, and metabolism in EA.hy926 endothelial cells, when given in a concentration-dependent and time-dependent manner.<br />\r\nAfter the treatment with serum increasing concentrations and the respective time periods, it was observed that not only does serum have no cytotoxic effect on the cells but also improves their viability. This determination can be supported by the significant increase in glucose uptake by the cells with an increase in serum concentrations. The maintenance of natural glucose homeostasis in the body depends on GLUT-4-mediated glucose transport in the cells [36]. As an increased amount of serum is provided for a prolonged time, the cells tend to make use of the high glucose environment which improves GLUT-4 expression and thereby reduces the insulin resistant condition which can support the increased glucose tolerance due to high GLUT-4 expression [37]. LDH also associated with endothelial-derived soluble adhesion molecules, recognized markers of endothelial activation that are commonly suppressed by NO [38]. From the results of the LDH assay, it can be understood that even though the cells produce higher amounts of LDH at increasing concentrations, their concentrations are not significant enough to establish that serum has an ameliorating effect on insulin resistance. The reduction in NO could be attributed to the condition of endothelial dysfunction thereby hindering vaso homeostasis. The conclusion drawn should further be confirmed with the help of gene expression studies that are done by targeting and understanding the specific factors involved like eNOS, and GLUT-4 to establish an appropriate hypothesis of the insulin resistance mediated endothelial dysfunction (Figure 5).<br />\r\n<a href=\"https://www.bsmiab.org/jabet/wp-content/uploads/sites/2/2022/07/178-1655357396.pdf\">Figure 5.</a> Effect of growth factors in serum on insulin resistance and endothelial dysfunction. In the ideal pathway, IRS is phosphorylated by Insulin, which activates the MAP Kinase signaling pathway for cell growth, proliferation and gene expression, as well as the PI3K signaling pathway for GLUT-4 translocation, lipid synthesis and cell proliferation. Insulin resistance results in the disruption of these signaling pathways. ET-1 is a vasoconstrictor, while NO is a vasodilator. Endothelial dysfunction occurs as a result of IR disrupting the balance between these levels by decreasing the NO levels and increasing the ET-1 levels.</p>"
},
{
"section_number": 6,
"section_title": "ACKNOWLEDGEMENT",
"body": "<p>None.</p>"
},
{
"section_number": 7,
"section_title": "AUTHORS CONTRIBUTION",
"body": "<p>HM, PT and AP drafted the manuscript. HM, PT, RK, AP and AB performed the experiments. AM proofread the manuscript and helped in analyzing the data. SS designed the experiments, analyzed, corrected, and validated the results. All authors read and approved the final submitted version of the manuscript.</p>"
},
{
"section_number": 8,
"section_title": "CONFLICTS OF INTEREST",
"body": "<p>There is no conflict of interest among the authors.</p>"
}
],
"figures": [
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/39/25/178-1655357396-Figure1.jpg",
"caption": "Figure 1. Assessment of cell viability by MTT in endothelial cells after treatment with fetal bovine serum in a time (5 min, 10min, 15min) and dose (5%, 10%, 15%) dependent manner. Triton X-100 (0.01%) is a positive control and DMSO serves as solvent control. Data represents the mean ± SEM of triplicates (N=3). #p < 0.001 as compared to control.",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/39/25/178-1655357396-Figure2.jpg",
"caption": "Figure 2. Nitric Oxide estimation in endothelial cells after treatment with fetal bovine serum in a time (5 min, 10min, 15min) and dose (5%, 10%, 15%) dependent manner, by using NED and sulphanilamide reagents. L-NAME is a positive control. Data represents the mean ± SEM of triplicates (N=3). *p < 0.05 and #p < 0.001 as compared to control.",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/39/25/178-1655357396-Figure3.jpg",
"caption": "Figure 3. LDH estimation in endothelial cells to observe cell cytotoxicity after treatment with fetal bovine serum in a time (5 min, 10min, 15min) and dose (5%, 10%, 15%) dependent manner. Triton X-100 (0.01%) is the positive control. Data represents the mean ± SEM of triplicates (N=3). #p < 0.001 as compared to control.",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/39/25/178-1655357396-Figure4.jpg",
"caption": "Figure 4. Estimation of percentage of glucose uptake by 2-NBDG which is a fluorescent analogue of glucose, in endothelial cells after treatment with fetal bovine serum in a time (5 min, 10min, 15min) and dose (5%, 10%, 15%) dependent manner. Insulin (100nM) is a positive control and DMSO (0.01%) serves as solvent control. Data represents the mean ± SEM of triplicates (N=3). *p < 0.05 as compared to control.",
"featured": false
}
],
"authors": [
{
"id": 500,
"affiliation": [
{
"affiliation": "Department of Biotechnology, School of Bioengineering, SRM Institute of Science and Technology, Kattankulathur-603 203, Tamil Nadu, India"
}
],
"first_name": "Harika",
"family_name": "Maganti",
"email": null,
"author_order": 1,
"ORCID": null,
"corresponding": false,
"co_first_author": true,
"co_author": false,
"corresponding_author_info": "",
"article": 125
},
{
"id": 501,
"affiliation": [
{
"affiliation": "Department of Biotechnology, School of Bioengineering, SRM Institute of Science and Technology, Kattankulathur-603 203, Tamil Nadu, India"
}
],
"first_name": "Prabu",
"family_name": "Thandapani",
"email": null,
"author_order": 2,
"ORCID": null,
"corresponding": false,
"co_first_author": true,
"co_author": false,
"corresponding_author_info": "",
"article": 125
},
{
"id": 502,
"affiliation": [
{
"affiliation": "Department of Biotechnology, School of Bioengineering, SRM Institute of Science and Technology, Kattankulathur-603 203, Tamil Nadu, India"
}
],
"first_name": "Ragulprasath",
"family_name": "Kailasam",
"email": null,
"author_order": 3,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 125
},
{
"id": 503,
"affiliation": [
{
"affiliation": "Department of Biotechnology, School of Bioengineering, SRM Institute of Science and Technology, Kattankulathur-603 203, Tamil Nadu, India"
}
],
"first_name": "Adithi",
"family_name": "Pisapati",
"email": null,
"author_order": 4,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 125
},
{
"id": 504,
"affiliation": [
{
"affiliation": "Department of Biotechnology, School of Bioengineering, SRM Institute of Science and Technology, Kattankulathur-603 203, Tamil Nadu, India"
}
],
"first_name": "Akshaya",
"family_name": "Bala",
"email": null,
"author_order": 5,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 125
},
{
"id": 505,
"affiliation": [
{
"affiliation": "Department of Biotechnology, School of Bioengineering, SRM Institute of Science and Technology, Kattankulathur-603 203, Tamil Nadu, India"
}
],
"first_name": "Angel",
"family_name": "Mendonca",
"email": null,
"author_order": 6,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 125
},
{
"id": 506,
"affiliation": [
{
"affiliation": "Department of Biotechnology, School of Bioengineering, SRM Institute of Science and Technology, Kattankulathur-603 203, Tamil Nadu, India"
}
],
"first_name": "Sujatha",
"family_name": "Sundaresan",
"email": "sujathas1@srmist.edu.in",
"author_order": 7,
"ORCID": "http://orcid.org/0000-0001-7676-2130",
"corresponding": true,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "Sujatha Sundaresan, PhD; Department of Biotechnology, School of Bioengineering, SRM Institute of Science and Technology, Kattankulathur, Tamil Nadu, India, e-mail: sujathas1@srmist.edu.in",
"article": 125
}
],
"views": 904,
"downloads": 186,
"references": [
{
"id": 4131,
"serial_number": 1,
"pmc": null,
"reference": "Behm B, Babilas P, Landthaler M, Schreml S. Cytokines, chemokines and growth factors in wound healing. Journal of the European Academy of Dermatology and Venereology 2012;26:812–20.",
"DOI": null,
"article": 125
},
{
"id": 4132,
"serial_number": 2,
"pmc": null,
"reference": "Wee P, Wang Z. Epidermal growth factor receptor cell proliferation signaling pathways. Cancers (Basel) 2017;9:1–45..",
"DOI": null,
"article": 125
},
{
"id": 4133,
"serial_number": 3,
"pmc": null,
"reference": "Subbiahanadar Chelladurai K, Selvan Christyraj JD, Rajagopalan K, Yesudhason BV, Venkatachalam S, Mohan M, et al. Alternative to FBS in animal cell culture – An overview and future perspective. Heliyon 2021;7:e07686.",
"DOI": null,
"article": 125
},
{
"id": 4134,
"serial_number": 4,
"pmc": null,
"reference": "Boucher J, Kleinridders A, Kahn CR. Insulin receptor signaling in normal and insulin-resistant states. Cold Spring Harbor perspectives in biology 2014;6: a009191.",
"DOI": null,
"article": 125
},
{
"id": 4135,
"serial_number": 5,
"pmc": null,
"reference": "Barac A, Campia U, Panza JA. Methods for evaluating endothelial function in humans. Hypertension 2007;49:748–60.",
"DOI": null,
"article": 125
},
{
"id": 4136,
"serial_number": 6,
"pmc": null,
"reference": "Król M, Kepinska M. Human nitric oxide synthase—its functions, polymorphisms, and inhibitors in the context of inflammation, diabetes and cardiovascular diseases. International Journal of Molecular Sciences 2021;22:1–18.",
"DOI": null,
"article": 125
},
{
"id": 4137,
"serial_number": 7,
"pmc": null,
"reference": "Sandoo A, Veldhuijzen van Zanten JJCS, Metsios GS, Carroll D, Kitas GD. The Endothelium and Its Role in Regulating Vascular Tone. The Open Cardiovascular Medicine Journal 2015;4:302–12.",
"DOI": null,
"article": 125
},
{
"id": 4138,
"serial_number": 8,
"pmc": null,
"reference": "Muniyappa R, Sowers JR. Role of insulin resistance in endothelial dysfunction. Reviews in Endocrine and Metabolic Disorders 2013;14:5–12.",
"DOI": null,
"article": 125
},
{
"id": 4139,
"serial_number": 9,
"pmc": null,
"reference": "Meza CA, la Favor JD, Kim DH, Hickner RC. Endothelial dysfunction: Is there a hyperglycemia-induced imbalance of NOX and NOS? International Journal of Molecular Sciences 2019;20.",
"DOI": null,
"article": 125
},
{
"id": 4140,
"serial_number": 10,
"pmc": null,
"reference": "Davis JA, Sharma S, Mittra S, Sujatha S, Kanaujia A, Shukla G, et al. Antihyperglycemic effect of Annona squamosa hexane extract in type 2 diabetes animal model: PTP1B inhibition, a possible mechanism of action. Indian Journal of Pharmacology 2012;44:326–32.",
"DOI": null,
"article": 125
},
{
"id": 4141,
"serial_number": 11,
"pmc": null,
"reference": "Bryan NS, Grisham MB. Methods to detect nitric oxide and its metabolites in biological samples. Free Radical Biology and Medicine 2007;43:645–57.",
"DOI": null,
"article": 125
},
{
"id": 4142,
"serial_number": 12,
"pmc": null,
"reference": "Thomas MG, Marwood RM, Parsons AE, Parsons RB. The effect of foetal bovine serum supplementation upon the lactate dehydrogenase cytotoxicity assay: Important considerations for in vitro toxicity analysis. Toxicology in Vitro 2015;30:300–8.",
"DOI": null,
"article": 125
},
{
"id": 4143,
"serial_number": 13,
"pmc": null,
"reference": "Blodgett AB, Kothinti RK, Kamyshko I, Petering DH, Kumar S, Tabatabai NM. A fluorescence method for measurement of glucose transport in kidney cells. Diabetes Technology and Therapeutics 2011;13:743–51. https://doi.org/10.1089/dia.2011.0041.",
"DOI": null,
"article": 125
},
{
"id": 4144,
"serial_number": 14,
"pmc": null,
"reference": "Su Y, Qadri SM, Wu L, Liu L. Methylglyoxal modulates endothelial nitric oxide synthase-associated functions in EA.hy926 endothelial cells. Cardiovascular Diabetology 2013;12:134.",
"DOI": null,
"article": 125
},
{
"id": 4145,
"serial_number": 15,
"pmc": null,
"reference": "Zou C, Wang Y, Shen Z. 2-NBDG as a fluorescent indicator for direct glucose uptake measurement. Journal of Biochemical and Biophysical Methods 2005;64:207–15.",
"DOI": null,
"article": 125
},
{
"id": 4146,
"serial_number": 16,
"pmc": null,
"reference": "Muniyappa R, Quon MJ. Is further research needed on glucosamine?. The Journal of family practice 2007;56:343.",
"DOI": null,
"article": 125
},
{
"id": 4147,
"serial_number": 17,
"pmc": null,
"reference": "Price CL, Knight SC. Methylglyoxal: possible link between hyperglycaemia and immune suppression? Trends in Endocrinology and Metabolism 2009;20:312–7.",
"DOI": null,
"article": 125
},
{
"id": 4148,
"serial_number": 18,
"pmc": null,
"reference": "Santos JH, Hunakova L, Chen Y, Bortner C, Van Houten B. Cell sorting experiments link persistent mitochondrial DNA damage with loss of mitochondrial membrane potential and apoptotic cell death. The Journal of Biological Chemistry 2003;278:1728–34.",
"DOI": null,
"article": 125
},
{
"id": 4149,
"serial_number": 19,
"pmc": null,
"reference": "di Loreto S, Zimmitti V, Sebastiani P, Cervelli C, Falone S, Amicarelli F. Methylglyoxal causes strong weakening of detoxifying capacity and apoptotic cell death in rat hippocampal neurons. International Journal of Biochemistry and Cell Biology 2008;40:245–57.",
"DOI": null,
"article": 125
},
{
"id": 4150,
"serial_number": 20,
"pmc": null,
"reference": "Chang T, Untereiner A, Liu J, Wu L. Interaction of methylglyoxal and hydrogen sulfide in rat vascular smooth muscle cells. Antioxidants & redox signaling 2010;12:1093–1100.",
"DOI": null,
"article": 125
},
{
"id": 4151,
"serial_number": 21,
"pmc": null,
"reference": "Kim HK, Ha SH, Han J. Potential therapeutic applications of tetrahydrobiopterin: From inherited hyperphenylalaninemia to mitochondrial diseases. Annals of the New York Academy of Sciences 2010;1201:177–82.",
"DOI": null,
"article": 125
},
{
"id": 4152,
"serial_number": 22,
"pmc": null,
"reference": "Abudukadier A, Fujita Y, Obara A, Ohashi A, Fukushima T, Sato Y, et al. Tetrahydrobiopterin Has a Glucose-Lowering Effect by Suppressing Hepatic Gluconeogenesis in an Endothelial Nitric Oxide Synthase-Dependent Manner in Diabetic Mice. Diabetes 2013;62:3033–43.",
"DOI": null,
"article": 125
},
{
"id": 4153,
"serial_number": 23,
"pmc": null,
"reference": "Okazaki T, Otani H, Shimazu T, Yoshioka K, Fujita M, Katano T, et al. Reversal of inducible nitric oxide synthase uncoupling unmasks tolerance to ischemia/reperfusion injury in the diabetic rat heart. Journal of molecular and cellular cardiology 2011;50:534–44.",
"DOI": null,
"article": 125
},
{
"id": 4154,
"serial_number": 24,
"pmc": null,
"reference": "Khoo JP, Zhao L, Alp NJ, Bendall JK, Nicoli T, Rockett K, et al. Pivotal Role for Endothelial Tetrahydrobiopterin in Pulmonary Hypertension. Circulation 2005;111:2126-33.",
"DOI": null,
"article": 125
},
{
"id": 4155,
"serial_number": 25,
"pmc": null,
"reference": "Foxton RH, Land JM, Simon AE, Heales JR. Tetrahydrobiopterin Availability in Parkinson’s and Alzheimer’s Disease; Potential Pathogenic Mechanisms. Neurochemical research 2007;32:751–756.",
"DOI": null,
"article": 125
},
{
"id": 4156,
"serial_number": 26,
"pmc": null,
"reference": "Cheng H, Wang H, Fan X, Paueksakon P, Harris RC. Improvement of endothelial nitric oxide synthase activity retards the progression of diabetic nephropathy in db/db mice. Kidney International 2012;82:1176–83.",
"DOI": null,
"article": 125
},
{
"id": 4157,
"serial_number": 27,
"pmc": null,
"reference": "Thida M, Earl J, Zhao Y, Wang H, Tse CS, Vickers JJ, et al. Effects of Sepiapterin Supplementation and NOS Inhibition on Glucocorticoid-Induced Hypertension. American journal of hypertension 2010;23:569–74.",
"DOI": null,
"article": 125
},
{
"id": 4158,
"serial_number": 28,
"pmc": null,
"reference": "Tiefenbacher CP, Lee CH, Kapitza J, Dietz V, Niroomand F. Sepiapterin reduces postischemic injury in the rat heart. European journal of physiology 2003;447:1–7.",
"DOI": null,
"article": 125
},
{
"id": 4159,
"serial_number": 29,
"pmc": null,
"reference": "Crabtree MJ, Brixey R, Batchelor H, Hale AB, Channon KM. Integrated redox sensor and effector functions for tetrahydrobiopterin- and glutathionylation-dependent endothelial nitric-oxide synthase uncoupling. Journal of Biological Chemistry 2013;288:561–9.",
"DOI": null,
"article": 125
},
{
"id": 4160,
"serial_number": 30,
"pmc": null,
"reference": "Luiking YC, Engelen MP, Deutz NE. Regulation of nitric oxide production in health and disease. Current Opinion in Clinical Nutrition and Metabolic Care 2010;13:97–104.",
"DOI": null,
"article": 125
},
{
"id": 4161,
"serial_number": 31,
"pmc": null,
"reference": "Wu G, Meininger CJ. Nitric oxide and vascular insulin resistance. BioFactors 2009;35:21–7.",
"DOI": null,
"article": 125
},
{
"id": 4162,
"serial_number": 32,
"pmc": null,
"reference": "Arnal JF, Münzel T, Venema RC, James NL, Bai CL, Mitch WE, et al. Interactions between L-arginine and L-glutamine change endothelial NO production: An effect independent of NO synthase substrate availability. Journal of Clinical Investigation 1995;95:2565–72.",
"DOI": null,
"article": 125
},
{
"id": 4163,
"serial_number": 33,
"pmc": null,
"reference": "Hadi HA, Carr CS, Al Suwaidi J. Endothelial dysfunction: cardiovascular risk factors, therapy, and outcome. Vascular health and risk management 2005;1:183–98.",
"DOI": null,
"article": 125
},
{
"id": 4164,
"serial_number": 34,
"pmc": null,
"reference": "Ainscow EK, Zhao C, Rutter GA. Acute overexpression of lactate dehydrogenase-A perturbs β-cell mitochondrial metasbolism and insulin secretion. Diabetes 2000;49:1149–55.",
"DOI": null,
"article": 125
},
{
"id": 4165,
"serial_number": 35,
"pmc": null,
"reference": "Kaja S, Payne AJ, Naumchuk Y, Koulen P. Quantification of lactate dehydrogenase for cell viability testing using cell lines and primary cultured astrocytes. Current Protocols in Toxicology 2017;72:1–10.",
"DOI": null,
"article": 125
},
{
"id": 4166,
"serial_number": 36,
"pmc": null,
"reference": "Zisman A, Peroni OD, Abel ED, Michael MD, Mauvais-Jarvis F, Lowell BB, et al. Targeted disruption of the glucose transporter 4 selectively in muscle causes insulin resistance and glucose intolerance. Nature Medicine 2000;6:924–28.",
"DOI": null,
"article": 125
},
{
"id": 4167,
"serial_number": 37,
"pmc": null,
"reference": "Wang T, Wang J, Hu X, Huang XJ, Chen GX. Current understanding of glucose transporter 4 expression and functional mechanisms. World Journal of Biological Chemistry 2020;11:76–98.",
"DOI": null,
"article": 125
},
{
"id": 4168,
"serial_number": 38,
"pmc": null,
"reference": "Kato GJ, McGowan V, Machado RF, Little JA, Taylor VI J, Morris CR, et al. Lactate dehydrogenase as a biomarker of hemolysis-associated nitric oxide resistance, priapism, leg ulceration, pulmonary hypertension, and death in patients with sickle cell disease. Blood 2006;107:2279–85.",
"DOI": null,
"article": 125
}
]
},
{
"id": 126,
"slug": "178-1654118747-evaluation-of-the-thrombolytic-activity-of-the-methanolic-extract-of-melia-azedarach-fruits-and-leaves-in-vitro",
"featured": false,
"slider": false,
"issue": "Vol5 Issue3",
"type": "original_article",
"manuscript_id": "178-1654118747",
"recieved": "2022-06-01",
"revised": null,
"accepted": "2022-07-01",
"published": "2022-07-26",
"pdf_file": "https://jabet.bsmiab.org/media/pdf_file/2023/28/178-1654118747.pdf",
"title": "Evaluation of the thrombolytic activity of the methanolic extract of Melia azedarach fruits and leaves in vitro",
"abstract": "<p><em>Melia azedarach</em> is a plant of the Meliaceae family, used worldwide in various medical fields. Because of the widespread incidence of thrombosis worldwide, especially during the coronavirus epidemic, this study was conducted to evaluate the in vitro thrombolytic effect of methanolic extracts of <em>Melia azedarach</em> leaves and fruits. Series of dilutions starting from 2 mg/ml to 20mg/ml were prepared from the methanolic extracts. The results showed that thrombolysis rates were between (18.7%-29.3%) for fruit extract in distilled water, (18.3%-30.1%) in phosphate buffer, (19.3%-35%) for leaves extract in distilled water and (20%-32.3%) in phosphate buffer for dilutions from 2 mg/ml to 20mg/ml. The streptokinase (positive control) had a thrombolytic effect of 47.54 % in distilled water and 44.36 % in phosphate buffer, compared to negative controls of 5.94 % and 6.34 %, respectively. Phytochemical screening found flavonoids and coumarins in leaves extract and only flavonoids in fruit extract, suggesting that the increase in thrombolytic effect may be attributed to these flavonoids and coumarins. The total phenolic content was 15.78 (mg GAE\\1g) in leaves extract and 3.64 (mg GAE\\1g) in fruit extract, while the total flavonoid content was 0.813 (mg QE\\1g) in leaves extract and 0.17 (mg QE\\1g) in fruit extract. In conclusion, these results showed that <em>Melia azedarach</em> has a thrombolytic effect.</p>",
"journal_reference": "J Adv Biotechnol Exp Ther. 2022; 5(3): 634-641.",
"academic_editor": "Akhi Moni, PhD; ABEx Bio-Research Center, Dhaka-1230, Bangladesh",
"cite_info": "Hussain HG, Agha MH. Evaluation of the thrombolytic activity of the methanolic extract of Melia azedarach fruits and leaves in vitro. J Adv Biotechnol Exp Ther. 2022; 5(3): 634-641.",
"keywords": [
"flavonoid",
"Methanolic extract",
"Melia azedarach",
"phenol",
"Thrombolytic effect"
],
"DOI": "doi.org/10.5455/jabet.2022.d142",
"sections": [
{
"section_number": 1,
"section_title": "INTRODUCTION",
"body": "<p>Coagulation is a complex physiological process performed to stop bleeding, where the blood clot is formed of fibrin, platelets, red and white blood cells [<a href=\"#r-1\">1</a>]. This happens when a blood vessel is damaged, and the platelets are activated, change their shape, then stick to the injury site and each other to create a plug that covers the damaged part and prevents blood from leaking [<a href=\"#r-2\">2</a>]. During this process, the dissolved fibrinogen transforms into undissolved fibrin and then intertwines with the platelets in the plug to create a network that traps more platelets and cells [<a href=\"#r-2\">2</a>]. But when dysfunction occurs in this process, it will lead to three possibilities: increasing the risk of bleeding, blood clotting, or increasing both. When a blood clot is formed, in this case, it is called thrombosis [<a href=\"#r-3\">3</a>].<br />\r\nInfectious diseases and respiratory infections were the leading causes of death worldwide until 1990 when cardiovascular diseases overtook them. After the coronavirus infection, respiratory disorders resurfaced as the leading cause of death, and the risk of thrombosis in respiratory patients has risen from 5% to 16% [<a href=\"#r-4\">4</a>]. The widely utilized thrombolytic agents have shown numerous drawbacks in recent years. These include alteplase, anistreplase, streptokinase (SK), urokinase (UK), and plasminogen activator (tPA), whereas the most commonly used agents are UK and SK [<a href=\"#r-5\">5</a>]. However, these drugs are not very safe due to the immunogenicity factors, high risk of bleeding, allergic reaction, and lack of specificity. Because of the deficiencies of existing thrombolysis agents, extensive research is conducted to find more effective drugs [<a href=\"#r-6\">6</a>].<br />\r\nPlants are the primary sources of natural compounds. According to some reports, approximately 30% of pharmaceuticals worldwide are derivatives from plants, whereas they are considered safer and less toxic in terms of side effects. So that more research is needed to find a new source of natural compounds that have thrombolytic effects [<a href=\"#r-5\">5</a>].<br />\r\n<em>Melia azedarach </em>is commonly known as the chinaberry tree, pride of India, bead tree, and Cape lilac. It is a kind of deciduous tree from the mahogany family, Meliaceae. It is a small to medium-sized tree or shrub, ranging in height from 5 to 15 meters. It is native to Asia and then moved to America, Africa, and Arab countries [<a href=\"#r-7\">7,8</a>].<br />\r\nAccording to previous studies, <em>Melia azedarach </em>contains many chemical compounds, including terpenes, steroids, alkaloids, flavonoids, anthraquinones, lignans, and coumarins [<a href=\"#r-9\">9</a>]. This plant has several effects: liver protective, anti-fertility, antioxidant, antipyretic, anti-viral, anti-bacterial, anti-nephrolithiasis, anti-cancer, anti-ulcer, anti-protozoal, and anti-parasitic effects, and helps heal wounds [<a href=\"#r-7\">7</a>].<br />\r\nThis study was conducted to evaluate the in vitro thrombolytic effect of methanolic extract of <em>Melia azedarach</em> fruits and leaves due to the increasing death rates as a result of thrombosis, particularly during the coronavirus pandemic, and the need for a new source of natural compounds that affect thrombosis effectively and without side effects.</p>"
},
{
"section_number": 2,
"section_title": "MATERIALS AND METHODS",
"body": "<p><strong>Plant collection and drying</strong><br />\r\nYellow ripe fruits and green leaves were collected in September from Damascus university garden, then were identified by Prof. Dr. Imad Kady, Department of Pharmacognosy and Medicinal Plant, Faculty of Pharmacy, Damascus University. The collected parts were dried in the shade for 3-4 days and then crushed.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Preparing the extracts</strong><br />\r\nA 100g of each dried fruit and leaves were combined with 250 ml methanol 99% and soaked for a week at room temperature. After a week, the extracts were filtered and dried at room temperature until their weight stabilized.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Phytochemical screening</strong><br />\r\nA preliminary phytochemical screening was performed on methanolic extracts of leaves and fruits to determine the presence of distinct chemical groups in the compounds. Then a qualitative screening was performed to determine flavonoids, alkaloids, coumarins, anthraquinones, and saponins [<a href=\"#r-10\">10</a>].</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Preparation of dilutions</strong><br />\r\nA 200 mg of each dried extract, fruits and leaves were dissolved in 10 ml of distilled water (DW) or phosphate buffer (PB) to prepare a solution with a concentration of 20 mg/ml. Phosphate buffer was used to have a pH similar to blood pH 7.4. A series of different dilutions (2-4-6-8-10-12-14-16-18-20) mg/ml was prepared for each extract. So that we had 4 standard series of dilutions: leaves extract in distilled water, leaves extract in phosphate buffer, fruits extract in distilled water, and fruits extract in phosphate buffer.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Preparation of streptokinase (SK)</strong><br />\r\nStreptokinase suspension of 100 μl (30,000 IU) was used in this study, prepared by adding 5 mL of distilled water or phosphate buffer to a commercially available lyophilized SK vial with a concentration of 1,500,000 IU [<a href=\"#r-11\">11</a>].</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Collection of the blood</strong><br />\r\nWhole blood was drawn from healthy human volunteers without a history of oral contraceptives or anticoagulant therapy. Every 1 ml of blood was transferred to a previously weighted sterile Eppendorf tube. The ethical approval number is (1-9-2020 ph. F-Damascus-Un). The informed consent was obtained orally.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Determination of thrombolytic activity</strong><br />\r\nEppendorf tubes were incubated at 37°C for 45 minutes. After thrombus (clots) formation, the serum was completely removed without destroying the thrombus. Then each tube was weighed again to determine the weight of the thrombus (Thrombus weight = weight of the tube containing the thrombus – weight of the empty tube). A 100 μl of extracts of each concentration previously prepared was added to Eppendorf tubes containing thrombus.<br />\r\nPositive control: 100 μl of prepared streptokinase in distilled water.<br />\r\n100 μl of prepared streptokinase in phosphate buffer.<br />\r\nNegative control: 100 μl of distilled water, 100μl of phosphate buffer.<br />\r\nAll tubes were incubated at 37 °C for 90 min. The fluid was removed from the tubes and weighed. The weight difference as a percentage was calculated.<br />\r\nThrombolysis percentage % = (the difference in the weight of the thrombus between before and after the addition of the extracts/weight of the thrombus before the addition) X 100 [<a href=\"#r-11\">11</a>].</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Determination of total phenolic content</strong><br />\r\nThe Folin-Ciocalteu method was used to evaluate the total phenol in methanolic extracts of the fruits and leaves. A 100 μl of Folin-Ciocalteu reagent was mixed with a 20 μl of extracts solution sample in a test tube. Then 1.58 ml of distilled water was added with shaking and incubation in the shade for 9 min at room temperature. Next, a 300 μl of sodium carbonate (20%) was added to the test tube that was kept in the shade for 45 min. The absorbance of all samples was measured at 765 nm with a UV-visible spectrophotometer, and the total phenolic content of extracts was expressed as mg of Gallic acid Equivalent (GAE) per 1g of dried plant. (Gallic acid was used as standard) [<a href=\"#r-12\">12</a>].</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Determination of total flavonoids content</strong><br />\r\nThe total flavonoid content of methanolic extracts for fruits and leaves was determined by the aluminium chloride colorimetric method. In the test tube, 1ml of aluminium chloride (2%) was added to 1ml of the sample. Then the tube was shaken and closed to incubate in the shade for 30 min. The absorbance of all samples was measured at 464 nm with UV-visible spectrophotometer and using methanol as a blank. The total flavonoid content in each sample was expressed in mg of quercetin equivalent per 1g of dried extracts (quercetin was used as standard) [<a href=\"#r-13\">13</a>].</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Statistical analysis</strong><br />\r\nThe statistical study was conducted using GraphPad Prism 8.0.1 statistical program. The numerical data were expressed as the mean and standard deviation and were evaluated by one-way analysis of variance (ANOVA).</p>"
},
{
"section_number": 3,
"section_title": "RESULTS",
"body": "<p><strong>The thrombolytic activity for leaves and fruit extracts</strong><br />\r\nIn DW, the fruits, and leaves extracts showed moderate thrombolytic activity: 29.34<u>+</u>0.20% for fruits extract and 35.03<u>+</u>0.38% for leaves extract, compared to streptokinase (47.54<u>+</u>0.26%). In PB, the thrombolytic effect was 30.15<u>+</u>0.6% for fruit extract and 32.32+1.6% for leaves extract compared to streptokinase’s (44.36<u>+</u>0.91%). All results showed in <a href=\"#figure1\">Figures 1a-d </a>and <a href=\"#figure2\">Figure 2</a>.<br />\r\nThere was no significant difference neither between leaves extract in DW and PB (P value = 0.069), nor between fruit extracts in DW and PB (P value = 0.5).<br />\r\nComparing the results of the four series, a significant difference was found only between leaf extract in DW and fruit extract in DW (P-value = 0.002) and a significant difference between leaf extract in DW and fruit extract in PB (P-value = 0.005). Whereas the extract of leaves in DW had the best thrombolytic effect, the remaining three series (leaves in PB, fruits in DW, and fruits in PB) were all close in effect, and no significant difference was observed between them.<br />\r\nMoreover, comparing the highest percentages of thrombolysis reached by each prepared series at a concentration of 20 mg/ml with the positive controls, streptokinase in distilled water or phosphate buffer, we found P-value < 0.001. Therefore, there is a significant difference, which means that the prepared concentrations could not reach the thrombolytic effect of streptokinase (<a href=\"#figure1\">Figure 1 a-d</a>).</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>The phytochemical screening</strong><br />\r\nThe phytochemical screening indicated the presence of coumarins and flavonoids in leaves extract and only flavonoids in fruit extract.</p>\r\n\r\n<div id=\"figure1\">\r\n<figure class=\"image\"><img alt=\"\" height=\"290\" src=\"/media/article_images/2023/29/26/178-1654118747-Figure1.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 1. </strong>The thrombolytic activity (%) according to concentration of fruit extract (a) and leaves extract (b) in distilled water (DW). The thrombolytic activity % according to concentration of fruit extract (c) and leaves extract (d) in phosphate buffer (PW). *** indicates P values<0.001 compared between the thrombolytic activity of SK and 20 mg/ml of each extract.</figcaption>\r\n</figure>\r\n</div>\r\n\r\n<div id=\"figure2\">\r\n<figure class=\"image\"><img alt=\"\" height=\"268\" src=\"/media/article_images/2023/29/26/178-1654118747-Figure2.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 2.</strong> Comparison between the highest thrombolytic effect for all extracts with positive and negative controls.</figcaption>\r\n</figure>\r\n</div>\r\n\r\n<p><strong>Total phenol and flavonoids content in leaves and fruits</strong><br />\r\nThe determination of total phenol was based on the calibration curve of gallic acid used as standard (<a href=\"#figure3\">Figure 3</a>), and of flavonoid was based on the calibration curve of quercetin (<a href=\"#figure4\">Figure 4</a>). It was found that the amount of phenol and flavonoid was higher in leaves extract than in fruit extract (<a href=\"#Table-1\">Table 1</a>).</p>\r\n\r\n<div id=\"Table-1\">\r\n<p><a href=\"https://jabet.bsmiab.org/table/178-1654118747-table1/\">Table-1</a><strong>Table 1.</strong> Total phenolic content, and total flavonoids content of <em>Melia azedarach</em> fruits and leaves.</p>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<div id=\"figure3\">\r\n<figure class=\"image\"><img alt=\"\" height=\"321\" src=\"/media/article_images/2023/29/26/178-1654118747-Figure3.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 3. </strong>Standard curve of gallic acid from which the concentration of phenols was calculated.</figcaption>\r\n</figure>\r\n</div>\r\n\r\n<div id=\"figure4\">\r\n<figure class=\"image\"><img alt=\"\" height=\"298\" src=\"/media/article_images/2023/29/26/178-1654118747-Figure4.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 4. </strong>Standard curve of quercetin from which the concentration of flavonoids was calculated.</figcaption>\r\n</figure>\r\n</div>"
},
{
"section_number": 4,
"section_title": "DISCUSSION",
"body": "<p>This study was conducted to evaluate the in vitro thrombolytic effect of methanolic extracts of <em>Melia azedarach</em> fruits and leaves as part of discovering new thrombolytic drugs.<br />\r\nThere were no differences between PB and DW extracts in thrombolytic effects, so they can be used as extracts to give the same effects.<br />\r\nThe phytochemical screening confirmed the presence of flavonoids in the methanolic extract of fruits and leaves, as well as coumarins in leaves extract. The total phenolic and flavonoid content was higher in leaves extract. These outcomes explain why leaves have a better thrombolytic effect than fruits. These results agreed with a study by Guguloth et al. 2021, which attributed the thrombolytic effect to the flavonoids [<a href=\"#r-13\">13</a>], and the suggested mechanism is weakening platelets adhesion [<a href=\"#r-1\">1</a>]. In addition, a study by Alamgeer et al. 2018 attributed the thrombolytic effect to coumarins, whereby coumarins inhibit calcium activity [<a href=\"#r-14\">14</a>].<br />\r\nFor total phenolic content, comparing our study with other studies, Munir et al. 2012 found that the content of phenols reached 82.54 mg GAE/1g and 66.89 mg GAE/1g in leaves and fruits, respectively. In comparison, the flavonoid content reached 16.99 mg QE/1g and 10.67 mg QE/1g in leaves and fruits, respectively [<a href=\"#r-15\">15</a>]. However, we found lower values in our study, which were 15.78 mg GAE/1g in leaves and 3.64 mg GAE/1g in fruits for phenols; 0.813 mg QE/1g in leaves and 0.17mgQE/1g in fruits for flavonoids.<br />\r\nMoreover, the results of Mwamatope et al. were close to ours, where the concentration of phenols and flavonoids in the leaves, respectively, reached 7.828 mg GAE/1g and 0.532mgQE/1g [<a href=\"#r-16\">16</a>].<br />\r\nThe differences between these values and ours may be due to different methods of extraction, polar stores, geographical locations in which the plant was studied, and the different harvesting periods, which affected the chemical content of the plant [<a href=\"#r-16\">16</a>].</p>"
},
{
"section_number": 5,
"section_title": "CONCLUSION",
"body": "<p>Based on the above results, it is clear that this plant may have significant implications in the thrombolytic field. The extracts can be used in water and blood pH without significant differences. In addition, the leaves extract gave the best thrombolytic effect among the prepared extracts, which may be attributed to flavonoids and coumarins.<br />\r\nThis study also indicated the possibility of developing novel drugs in the thrombolytic field from <em>Melia azedarach</em>. This is only a preliminary study, and the extract should be thoroughly investigated phytochemically and pharmacologically to confirm its medicinal and pharmaceutical potential.</p>"
},
{
"section_number": 6,
"section_title": "ACKNOWLEDGEMENTS",
"body": "<p>This work was supported by Faculty of Pharmacy, Damascus University.</p>"
},
{
"section_number": 7,
"section_title": "AUTHORS CONTRIBUTION",
"body": "<p>MIHA supervised the overall research work, revised the manuscript. critically reviewed the manuscript. HH performed the research work, wrote the manuscript and analyzed the data, revised the manuscript. All authors revised and approved the final version of the manuscript.</p>"
},
{
"section_number": 8,
"section_title": "CONFLICTS OF INTEREST",
"body": "<p>There is no conflict of interest among the authors.</p>"
}
],
"figures": [
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/29/26/178-1654118747-Figure1.jpg",
"caption": "Figure 1. The thrombolytic activity (%) according to concentration of fruit extract (a) and leaves extract (b) in distilled water (DW). The thrombolytic activity % according to concentration of fruit extract (c) and leaves extract (d) in phosphate buffer (PW). *** indicates P values<0.001 compared between the thrombolytic activity of SK and 20 mg/ml of each extract.",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/29/26/178-1654118747-Figure2.jpg",
"caption": "Figure 2 Comparison between the highest thrombolytic effect for all extracts with positive and negative controls.",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/29/26/178-1654118747-Figure3.jpg",
"caption": "Figure 3. Standard curve of gallic acid from which the concentration of phenols was calculated.",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/29/26/178-1654118747-Figure4.jpg",
"caption": "Figure 4. Standard curve of quercetin from which the concentration of flavonoids was calculated.",
"featured": false
}
],
"authors": [
{
"id": 507,
"affiliation": [
{
"affiliation": "Department of pharmacognosy and medicinal plant, Faculty of pharmacy, Damascus University, Damascus, Syria."
}
],
"first_name": "Hiba Ghanem",
"family_name": "Hussain",
"email": null,
"author_order": 1,
"ORCID": "http://hiba56304@gmail.com",
"corresponding": true,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "Hiba Ghanem Hussain, Department of pharmacognosy and medicinal plant, Faculty of pharmacy, Damascus University, Damascus, Syria, e-mail: hiba56304@gmail.com",
"article": 126
},
{
"id": 508,
"affiliation": [
{
"affiliation": "Department of pharmacognosy and medicinal plant, Faculty of pharmacy, Damascus University, Damascus, Syria."
}
],
"first_name": "Mohammed Isam Hasan",
"family_name": "Agha",
"email": null,
"author_order": 2,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 126
}
],
"views": 878,
"downloads": 209,
"references": [
{
"id": 4169,
"serial_number": 1,
"pmc": null,
"reference": "Lichota A, Szewczyk EM, Gwozdzinski K. Factors affecting the formation and treatment of thrombosis by natural and synthetic compounds. Int J Mol Sci. 2020; 21: (1–54).",
"DOI": null,
"article": 126
},
{
"id": 4170,
"serial_number": 2,
"pmc": null,
"reference": "Furie B, Furie BC. In vivo thrombus formation. J Thromb Haemost. 2007; 5: (12–7).",
"DOI": null,
"article": 126
},
{
"id": 4171,
"serial_number": 3,
"pmc": null,
"reference": "Mackman N. Triggers, targets, and treatments for thrombosis. NIH. 2008; 451: (914–918).",
"DOI": null,
"article": 126
},
{
"id": 4172,
"serial_number": 4,
"pmc": null,
"reference": "Smilowitz NR, Subashchandran V, Yuriditsky E, Horowitz JM, Reynolds HR, Hochman JS, et al. Thrombosis in hospitalized patients with viral respiratory infections versus COVID-19. Am Heart J. 2021; 231: (93–95).",
"DOI": null,
"article": 126
},
{
"id": 4173,
"serial_number": 5,
"pmc": null,
"reference": "Sayeed MA, Kabir H, Mamun M, Rashid U, Bhuiyan FA, Rashid MA. Thrombolytic activity of methanolic extracts of Desmodium paniculatum (L.) and Sarcochlamys pulcherrima (Roxb.). Bangladesh Pharm J. 2014; 17: (67–69).",
"DOI": null,
"article": 126
},
{
"id": 4174,
"serial_number": 6,
"pmc": null,
"reference": "Sherwani SK, Bashir A, Haider SS, Shah MA. Thrombolytic potential of aqueous and methanolic crude extracts of Camellia sinensis (Green Tea): In vitro study. J Pharmacogn Phytochem. 2013; 2: (125-129).",
"DOI": null,
"article": 126
},
{
"id": 4175,
"serial_number": 7,
"pmc": null,
"reference": "Sultana S, Asif HM, Akhtar N, Waqas M, Rehman SU. Comprehensive review on ethanobotanical uses, phytochemistry and pharmacological properties of melia azedarachlinn. AJPRHC. 2014; 6: (1-7).",
"DOI": null,
"article": 126
},
{
"id": 4176,
"serial_number": 8,
"pmc": null,
"reference": "Nadeeka N N A, Kim M S, Chathuranga K, Ma J Y, Kim H, Lee JS. Melia azedarach extract exhibits a broad spectrum of antiviral effect in vitro and in vivo. J Biomed Transl Res 2020;21:125-136..Wang W, Xia Z, Tian Z, Jiang H, Zhan Y, Liu C, et al. Chemical constituents from the fruits of Melia azedarach (Meliaceae). Biochem Syst Ecol. 2020; 92: (94-104).",
"DOI": null,
"article": 126
},
{
"id": 4177,
"serial_number": 9,
"pmc": null,
"reference": "Agha MI, Monjed H. Pharmacognosy chemistry. Damascus University; 1997.",
"DOI": null,
"article": 126
},
{
"id": 4178,
"serial_number": 10,
"pmc": null,
"reference": "Tabassum F, Chadni SH, Mou KN, Hasif KI, Ahmed T, Akter M. In-vitro thrombolytic activity and phytochemical evaluation of leaf extracts of four medicinal plants of Asteraceae family. J Pharmacogn Phytochem. 2017; 6: (1166-1169).",
"DOI": null,
"article": 126
},
{
"id": 4179,
"serial_number": 11,
"pmc": null,
"reference": "Duraisankar M, Ravichandran V. Antipyretic potential of polyherbal ayurvedic products. Asian J Pharm Clin Res. 2012; 5: (146–150).",
"DOI": null,
"article": 126
},
{
"id": 4180,
"serial_number": 12,
"pmc": null,
"reference": "Katib R, Sultan N. Evaluation of total phenolic content, total flavonoids content and free radical scavenging activity of lepidium sativum l. seeds and leaves. Bull Pharm Sci. 2021; 44: (377–385).",
"DOI": null,
"article": 126
},
{
"id": 4181,
"serial_number": 13,
"pmc": null,
"reference": "Guguloth SK, Malothu N, DSNBK P. GC-MS analysis and thrombolytic property of methanolic leaf extracts of Terminalia pallida Brandis against carrageenan instigated tail thrombosis Model in Mice. Pharmacognosy Res. 2021; 14: (53–60).",
"DOI": null,
"article": 126
},
{
"id": 4182,
"serial_number": 14,
"pmc": null,
"reference": "Alamgeer A, Tarar M, Hasan UH, Saleem M. Evaluation of anticoagulant and thrombolytic activity of Berberis orthobotrys in animal model. Bangladesh J Pharmacol. 2018; 13: (196–202).",
"DOI": null,
"article": 126
},
{
"id": 4183,
"serial_number": 15,
"pmc": null,
"reference": "Munir A, Sultana B, Babar T, Bashir A, Amjad M, Hassan Q. Investigation on the antioxidant activity of leaves, fruit and stem bark of Dhraik (Melia azedarach). Eur J Appl Sci. 2012; 4: (47–5).",
"DOI": null,
"article": 126
},
{
"id": 4184,
"serial_number": 16,
"pmc": null,
"reference": "Mwamatope B, Tembo D, Chikowe I, Kampira E, Nyirenda C. Total phenolic contents and antioxidant activity of Senna singueana, Melia azedarach, Moringa oleifera and Lannea discolor herbal plants. Sci African . 2020; 9: e00481.",
"DOI": null,
"article": 126
}
]
},
{
"id": 127,
"slug": "178-1652146042-efficacy-of-soil-borne-burkholderia-cepacia-like-mb-01-strain-for-shobicron-and-vertimec-degradation",
"featured": false,
"slider": false,
"issue": "Vol5 Issue3",
"type": "original_article",
"manuscript_id": "178-1652146042",
"recieved": "2022-05-10",
"revised": null,
"accepted": "2022-07-05",
"published": "2022-07-26",
"pdf_file": "https://jabet.bsmiab.org/media/pdf_file/2023/57/178-1652146042.pdf",
"title": "Efficacy of soil-borne Burkholderia Cepacia-like MB-01 strain for shobicron and vertimec degradation",
"abstract": "<p>Pesticides are necessary for agriculture, yet their highly toxic ingredients harm the ecosystem. Due to their toxicity, uncontrolled releases of large quantities of pesticides pollute the environment and provide a larger health risk to plants, animals, and humans. Bacteria are capable of degrading such pollutants and saving our ecosystem. In this study, a bacterial strain was isolated from Shobicron and Vertimec-treated lady’s finger soil using enrichment culture. The strain was identified as <em>Burkholderia Cepacia</em>-Like MB-01 based on morphological, physiological, and biochemical traits, as well as phylogenetic analysis of the 16S rRNA sequence. The bacterium grew best at 35 °C with a pH of 7. Furthermore, it was susceptible (S) to Cefepime and Penicillin but intermediate (I) resistant to Carbapenem and Tetracycline and resistant (R) to Ciprofloxacin, Kanamycin, and Gentamycin in an antibiotic sensitivity test. The rate of shobicron and vertimec degradation was measured over a five-day period using Mineral Salt (MS) medium. In its optimum growth condition, shobicron and vertimec degradation rates were around 76 % and 80 %, respectively. The isolated bacterial strain was capable of detoxifying shobicron and vertimec in the experiment. As a result, the bacterial strain could exploit as a possible shobicron and vertimec degrader for pesticide bioremediation.</p>",
"journal_reference": "J Adv Biotechnol Exp Ther. 2022; 5(3): 642-653.",
"academic_editor": "Md. Masudur Rahman, PhD; Sylhet Agricultural University, Bangladesh",
"cite_info": "Moniruzzaman M, Jinnah MM, et al. Efficacy of soil-borne Burkholderia Cepacia-like MB-01 strain for shobicron and vertimec degradation. J Adv Biotechnol Exp Ther. 2022; 5(3): 642-653.",
"keywords": [
"Bacteria",
"Antibiotic sensitivity",
"Shobicron",
"Vertimec",
"Degradation"
],
"DOI": "10.5455/jabet.2022.d143",
"sections": [
{
"section_number": 1,
"section_title": "INTRODUCTION",
"body": "<p>Bioremediation is a procedure that uses microorganisms or their enzymes to treat polluted places in order to restore them to their previous state [<a href=\"#r-1\">1</a>]. Indeed, bioremediation is the act of breaking down harmful pollutants into less dangerous compounds using biological organisms. The bioremediation method involves a variety of live species. Majority of them are bacteria and fungus due to their quick growth rate and capacity to be genetically modified. To break down a pollutant, a widely used strategy for bioremediation involves activating naturally occurring microbial communities and providing them with nutrients and other needs. We should utilize bioremediation to clean soil, water, and other materials since it is less damaging and poses fewer risks to the environment [<a href=\"#r-2\">2</a>].<br />\r\nPesticides are a group of poisonous compounds used mostly in agriculture. A pesticide is a mixture of substances used to eliminate or control a pest, which can include vectors of human or animal disease, undesirable plant species, or animals that cause damage to agricultural food products, timber and timber products, or animal feed during production, processing, storage, or marketing. Thousands of pesticide formulations exist around the world, containing almost 1500 registered ingredients [<a href=\"#r-3\">3</a>]. More than 55 % of the area used for agricultural production in developing countries uses about 26 % of pesticides produced worldwide [<a href=\"#r-4\">4</a>]. Farmers in Bangladesh frequently use various pesticide models on their agricultural areas, typically on the advice of unskilled vendors [<a href=\"#r-5\">5</a>]. Pesticides are applied in granules, liquids, and powders. Depending on the attacking pests in Bangladesh, farmers have utilized pesticides such as cypermthrin, carbofuran, amcozinon, profenofos, abamectin, and malathion.<br />\r\nHazardous pesticides can have acute and/or chronic poisoning effects, putting children at danger, and are recognized as a worldwide concern. In many places of the world, their widespread use has resulted in health concerns and deaths, typically as a result of occupational exposure and unintentional poisonings. Agricultural and public health professionals are exposed to the most dangerous pesticides due to their handling, mixing, and application. Pesticide residues in food and, possibly, drinking water can be a concern for the general public [<a href=\"#r-6\">6</a>].<br />\r\nAgricultural, residential, and industrial wastes are the main contributors of water contamination. When sewage is dumped into rivers, it is the most contaminant of fresh water. The amount of dissolved oxygen in the water decreases, and the Biological Oxygen Demand (BOD) decreases. When large amounts of fertilizer, herbicides, and pesticides are washed into rivers by rain, they pose a major threat to human life [<a href=\"#r-7\">7</a>]. Shobicron is a brand name for two ingredients: cypermethrin and profenofos. It’s used to fight pests like fruit flies, white flies, and dragonflies in a variety of agricultural crops like brinjal, lady’s finger, cotton, guava, mango, and banana. Cypermethrin is a synthetic pyrethroid that is widely used as a pesticide in agriculture, household, and animal husbandry, primarily to treat cracks, crevices, and spots for insect control [<a href=\"#r-8\">8</a>]. Cypermethrin has a very poor water solubility [<a href=\"#r-9\">9</a>]. It moves from a liquid solution to an adjourned molecule because it is hydrophobic [<a href=\"#r-10\">10</a>]. The urine excretion of cypermethrin metabolites in humans was completed 48 hours after the last of five daily 1.5 mg doses [<a href=\"#r-11\">11</a>]. Profenofos is an insecticide containing organophosphates. It’s a pale yellow to amber-colored liquid with a garlic-like odor [<a href=\"#r-12\">12</a>]. The resistance of the acetylcholinesterase enzyme is the mechanism of action of profenofos. Although it is utilized as a racemate, the S (-) isomer is a far more powerful inhibitor. Profenofos can prevent the cholinesterase enzyme from working properly. This can lead the neurological system to become over stimulated, resulting in nausea, dizziness, and confusion. Higher doses of profenofos can cause even more severe side effects, including respiratory paralysis and death. Profenofos has also been shown to affect bees and fish. Vertimec, as a brand name, is a popular insecticide in Bangladesh, where it is used on a wide range of crops. Abamectin is the active component. It is employed in a variety of agricultural crops, including brinjal, lady’s finger, tea, jujube, litchi, and others, to combat pests such as red spinder mite and mite. Abamectin is widely used as an insecticide in agriculture and as an anti-parasitic medication in veterinary medicine, and it has resulted in significant environmental contamination by posing a risk to non-target soil invertebrates and aquatic systems. Avermectin is made up of two homologues: avermectin B1a and avermectin B1b [<a href=\"#r-13\">13</a>]. Abamectin is extremely poisonous to aquatic invertebrates and highly toxic to fish. As a result, abamectin should not be disposed of in water. Abamectin contaminates groundwater and harms the environment.<br />\r\nThus, the present study was carried out to isolate and characterize shobicron and vertimec degrading bacteria from lady’s finger field soil, as well as determine their degradation efficiency for further exploitation of the strain for potential bioaugmentation.</p>"
},
{
"section_number": 2,
"section_title": "MATERIALS AND METHODS",
"body": "<p><strong>Sample collection</strong><br />\r\nOne gram of shobicron and vertimec-treated soil sample was collected from agricultural fields in Budpara and Rajshahi, Bangladesh that had a 5-7-years history of shobicron and vertimec use. Soil sample was collected and employed as a source of inoculums for enrichment culture. This research work was carried out over the course of seven months, from May to December 2019. The entire project was carried out in the microbiology laboratory, Department of Genetic Engineering and Biotechnology of the University of Rajshahi, Bangladesh.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Cytotoxicity analysis of shobicron and vertimec using <em>Artemia salina</em> through LC50</strong><br />\r\nThe brine shrimp lethality bioassay is a useful technique for toxicity screening, heavy metal detection, pesticide detection, fungal toxins detection, and dental cytotoxicity testing [<a href=\"#r-14\">14</a>]. Brine shrimp eggs were cultured in artificial seawater to obtain nauplii. The nauplii were counted and placed in test tubes visually. Then, in three replications, different amounts of shobicron and vertimec were delivered to tubes, with control doses kept at the same level as before. To assess the percentage of survived nauplii after 12, 24, 36, and 48 hours, data was collected and statistically analyzed using probit mortality software.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Enrichment of shobicron and vertimec degrading bacteria</strong><br />\r\nOne gram of shobicron and vertimec-treated soil was suspended in 100 mL of mineral salts (MS) medium (Himedia, India) supplemented with shobicron and vertimec in separate 250 mL Erlenmayer flasks (Eisco lab, India). To account for potential abiotic disappearance of shobicron and vertimec, control flasks without an inoculum were also prepared. Shaking at 120rpm for 3 days on an orbital shaker was the principal enrichment method. The cultures were employed as a source of inoculum in the following tests once they acquired sufficient turbidity.<br />\r\nBy plating out on mineral salts agar medium containing shobicron and vertimec, microorganisms capable of decomposing shobicron and vertimec were identified from enrichment cultures. The plates were incubated at 37 °C for 2 to 3 days, and one strain that grew on the medium and had a greater degrading ability was named MB-01 and stored for future use.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Morphological, and biochemical characterization of the isolate</strong><br />\r\nAfter 24 hours of growth on MS agar at 37 °C, the colony features of the isolated bacteria (colony form, color, margin, opacity, consistency, elevation) were studied under an inverted microscope (LABOMED CxL). The morphological (Gram staining and motility tests) and biochemical (catalase, oxidase, TSI, methyl red, lactose fermentation tests, carbohydrates such as glucose, mannose, arabinose, sucrose, lactose, and cellulose intake) characterizations were carried out according to the protocol [<a href=\"#r-15\">15</a>].</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Antibiotic sensitivity test</strong><br />\r\nThe antibiotic sensitivity and resistance pattern of the isolated bacteria were determined using the Kirby- Bauer disc diffusion method [<a href=\"#r-16\">16</a>]. For the antibiotic sensitivity test, the isolated bacterial strain was cultured overnight in nutrient broths using a shaker at 37 °C and 160 rpm. The LB (Luria Bertani) agar medium (Himedia, India) was prepared first, and then the sterile liquid medium was dispersed in sterile petridishes for preparing culture plates. Each petridish received approximately (15-20) ml of the media and was placed in the laminar airflow cabinet to solidify. Then, for the isolate, overnight grown LB culture (OD = 0.5) was poured into the nutrient agar plate and dried. Antibiotic disks (Table 1) were prepared using commercially available and regularly prescribed antibiotics, which were inserted in the center of the plates and incubated overnight at 37 °C. After an overnight incubation period, the zones on the plate were observed and quantified using a millimeter scale.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Effect of pH and temperatures on bacterial growth</strong><br />\r\nThe media were prepared, and pH adjusted to a range of 5 to 8 on a scale of 1 to 8. The bacteria were then inoculated in a medium that had already been prepared in front of the laminar airflow cabinet. After the culture was inoculated, it was placed in an incubator to maintain the various temperature ranging from 25 °C to 45 °C with 5 °C interval to test their growth efficiency. By using a spectrophotometer (Wincom, China) to measure optical density at 600nm, the growth rates were recorded over time.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>PCR amplification and sequencing of 16S rRNA gene and phylogenetic analysis</strong><br />\r\nThe Tiangen DNA extraction kit was used to extract the genomic DNA of the isolated shobicron and vertimec degrading bacterium. The 16S rRNA genes were amplified using forward primer in a polymerase chain reaction (PCR) (27 F – AGA GTT TGA TCM TGG CTC AG and 1492 R – GGT TAC CTT GTT ACG ACT T). Invitrogen (Thermo fisher scientific, Bangladesh) provided all the PCR reagents. The PCR reactions were carried out in a thermal cycler (Applied Biosystem 9700) with the following amplification conditions: initial denaturation at 95 °C for 5 minutes, followed by 30 cycles of denaturation at 94 °C for 1 minute, annealing at 55 °C for 1 minute, extension at 72 °C for 1 minute, and final extension at 72 °C for 10 minutes. The Freeze ‘N Squeeze DNA Gel Extraction Spin Columns (Bio-Rad, India) were used to purify the PCR results, which were then sequenced on both strands on the genetic analyzer (Prism 310). Chromas, a bioinformatics program, were used to alter the sequences. The BLASTN (http://www.ncbi.nih.gov/BLAST) algorithm was used to compare the 16S rRNA gene sequences to the 16S rRNA gene sequences of other organisms. NCBI (National center for biotechnology information) BLAST (www.ncbi.nlm.nih.gov/Blast) was used to create the phylogenetic tree.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Determination of shobicorn and vertimec degradation rate</strong><br />\r\nThe isolate was grown in a 150 mL conical flask with 100 mL Mineral salts medium (MSM) supplemented with shobicron and vertimec to test its growth. A spectrophotometer was used to calculate the ability to the isolate to degrade pesticides by measuring turbidity at 600 nm. To compare the deterioration of the isolate, a control flask was utilized without pesticides. Control and isolates had their ideal density values evaluated and recorded.</p>"
},
{
"section_number": 3,
"section_title": "RESULTS",
"body": "<p><strong>Cytotoxicity test of the 99% pure shobicron and vertimec</strong><br />\r\nThe toxic effects of pure shobicron and vertimec on <em>A. salina</em> were investigated using a brine shrimp lethality assay. After only 12 hours of exposure, the pure shobicron and vertimec LC<sub>50</sub> were 10.102 g/ml and 9.871 g/ml, respectively, according to probit analysis, and no live nauplii remained after 48 hours (<a href=\"#Table-1\">Table 1</a>). Shobicron and vertimec exhibited a significant mortality effect on nauplii, as evidenced by visual detection.</p>\r\n\r\n<div id=\"Table-1\">\r\n<p><a href=\"https://jabet.bsmiab.org/table/178-1652146042-table1/\">Table-1</a><strong>Table 1. </strong>Determination of LC<sub>50</sub> value of the shobicron and vertimec against <em>Artemia salina</em> after 12, 24, 36, and 48 h of exposure.</p>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Isolation and screening of shobicron and vertimec degrading bacteria</strong><br />\r\nThe bacteria that degrade shobicron and vertimec were identified using 99% pure shobicron and vertimec at 2 µg/ml and 4 µg/ml, respectively. The shobicron and vertimec-enriched MS plates were incubated at 37 °C for 24 hours, and bacterial colonies were observed to develop on the medium containing the aforesaid concentrations of shobicron and vertimec (<a href=\"#figure1\">Figure 1a</a>), with no growth at concentrations higher than that.</p>\r\n\r\n<div id=\"figure1\">\r\n<figure class=\"image\"><img alt=\"\" height=\"218\" src=\"/media/article_images/2023/57/26/178-1652146042-Figure1.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 1. </strong>Isolated shobicrn and vertimec degrading bacterial strain. (a) Bacterial growth on MS agar plate supplemented with 4 µg/ml of shobicron. (b) Gram staining isolated strain indicating gram-negative and rod-shaped structure.</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Morphological and biochemical characterization</strong><br />\r\nThe strain formed a smooth colony with an average diameter of 0.6–1 mm in biochemical tests. The cells were straight rods with rounded ends, measuring 0.4–0.7 lm in width and 1.2–1.7 lm in length. It was a gram-negative bacterium (<a href=\"#figure1\">Figure 1b</a>). The strain was positive for oxidase, nitrate reduction, and beta-glactocidase, but not for arginine dihydrolase, beta-glucosidase (aesculin hydrolysis), protease (gelatin liquefaction), tryptophanase, or urease activity. L-arabinose, D-glucose, D-mannose, and D-mannitol were among the carbohydrates that the bacterium could use.<br />\r\nTo identify the isolated bacterium, the 16S rRNA gene was amplified by PCR and sequenced from Invent technologies, Bangladesh. The strain MB-01 demonstrated very high similarity with the species of genus <em>Burkholderia,</em> and more than 99% gene homology was observed with <em>Burkholderia cepecia</em> complex (Bcc) species. Phylogenetic analysis of 16S rRNA gene also clustered the strain GB-01 within the clade of <em>Burkholderia cepecia</em> complex (Bcc) species in phylogenetic tree (<a href=\"#figure2\">Figure 2</a>). On the basis of morphological, physiological and biochemical properties, combined with 16S rRNA gene sequence analysis, the strain MB-01 was identified as <em>Burkholderia cepacia</em>-like species.</p>\r\n\r\n<div id=\"figure2\">\r\n<figure class=\"image\"><img alt=\"\" height=\"331\" src=\"/media/article_images/2023/57/26/178-1652146042-Figure2.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 2. </strong>Phylogenetic tree analysis with similar bacterial strains. The tree was constructed by the neighbor-joining method based on 16S rRNA gene sequences of strain MB-01 and related <em>Burkholderia</em> type strain.</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Physiological characterization</strong><br />\r\nThis test was carried out to assess the optimum pH and temperature for bacterial growth. The strain was grown on MS media at temperatures of 25 °C, 30 °C, 35 °C, 40 °C, and 45 °C in a pH range of 5 to 9. After measuring OD at 600nm in a Spectrophotometer, it was observed that the strain’s optimum growth was best at pH 7 (<a href=\"#figure3\">Figure 3a</a>) and 35 °C (<a href=\"#figure3\">Figure 3b</a>).</p>\r\n\r\n<div id=\"figure3\">\r\n<figure class=\"image\"><img alt=\"\" height=\"220\" src=\"/media/article_images/2023/57/26/178-1652146042-Figure3.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 3. </strong>Optimization of pH and temperature. (a) Effect of pH on growth of the strain MB-01, where the maximum growth was obtained at pH 7; (b) Effect of temperature on bacterial growth, where maximum growth was observed at 35 °C. Data were recorded after 12 hours of incubation.</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Antibiotic sensitivity test</strong><br />\r\nThe result showed that <em>Burkholderia Cepacia</em>-Like MB-01 was susceptible (S) to Cefepime and Penicillin but intermediate (I) resistant to Carbapenem, and Tetracycline and resistant (R) to Ciprofloxacin, Kanamycin, and Gentamycin (<a href=\"#figure4\">Figure 4</a>).</p>\r\n\r\n<div id=\"figure4\">\r\n<figure class=\"image\"><img alt=\"\" height=\"272\" src=\"/media/article_images/2023/57/26/178-1652146042-Figure4.jpg\" width=\"253\" />\r\n<figcaption><strong>Figure 4. </strong>Antibiotic sensitivity test of the strain MB-01. Isolate was resistant to most of the tested antibiotics, and susceptible to Cefepime, and Penicillin. Note: Resistant=<10 mm; Intermediate resistance=10-15 mm; Susceptible=>15 mm.</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Shobicron and vertimec degrading efficiency of </strong><strong><em>Burkholderia Cepacia</em></strong><strong>-like MB-01</strong><br />\r\nThe strain MB-01 was cultivated on shobicron and vertimec supplemented liquid Mineral salts medium (MSM) at 35 °C for 5 days in order to gain relative degradation. The strain decomposed shobicron and vertimec, as evidenced by a steady increase in turbidity (OD600) over a 5-day period (<a href=\"#figure5\">Figure 5</a>). When shobicron and vertimec were employed as the only source of carbon, the strain’s growth was dramatically boosted when compared to the control. Over 5 days of incubation at 35 °C, strain MB-01 showed a clear increase in growth using shobicron and vertimec, with a 76 %, and a 80 % rise in OD<sub>600</sub>, respectively.<br />\r\nTo be more specific, we ran this experiment at different pH and temperature levels to see how well it degraded, and we obtained comparable results (<a href=\"#figure6\">Figure 6</a>) as shown in <a href=\"#figure5\">Figure 5</a>.</p>\r\n\r\n<div id=\"figure5\">\r\n<figure class=\"image\"><img alt=\"\" height=\"227\" src=\"/media/article_images/2023/57/26/178-1652146042-Figure5.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 5.</strong> Showing of degradation of shobicron (a) and vertimec (b) by the strain MB-01 in its optimum conditions. Data were recoded up to 5 days of incubation.</figcaption>\r\n</figure>\r\n</div>\r\n\r\n<div id=\"figure6\">\r\n<figure class=\"image\"><img alt=\"\" height=\"224\" src=\"/media/article_images/2023/57/26/178-1652146042-Figure6.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 6. </strong>Effect of pH (a) and temperature (b) on shobicron and vertimec degradation by the strain MB-01. Data were recorded after 5 days of incubation.</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>"
},
{
"section_number": 4,
"section_title": "DISCUSSION",
"body": "<p>Biological processes like microbial decomposition are considered a safe and effective way to remove contaminants from the environment [<a href=\"#r-17\">17, 18</a>]. Bioremediation is closely linked to the decomposition, or detoxification, of numerous pollutants and harmful compounds in the environment [<a href=\"#r-19\">19, 20</a>]. Some pesticides are quickly degraded by microorganisms, while others are harmful. Pesticides are metabolized by a variety of bacteria, including members of the genera <em>Alcaligenes, Flavobacterium, Pseudomonas</em>, and<em> Rhodococcus</em>. Microbial degradation is influenced by a wide range of environmental conditions in addition to the presence of bacteria. Pesticide degradation is influenced by a variety of environmental parameters such as pH, temperature, and nutrients [<a href=\"#r-21\">21, 22</a>]. Pesticides can be broken down by microorganisms that use them as carbon and energy sources in specific ways [<a href=\"#r-23\">23</a>].<br />\r\nBioremediation is a critical method for reducing pollution and restoring contaminated soil and water. We should utilize bioremediation to clean soil, water, and other materials since it is less damaging and poses fewer risks to the environment [<a href=\"#r-24\">24</a>]. Because of its environmental stability, toxicity, and biological reservation in organisms, pesticides cause serious health problems. Due to a lack of sufficient understanding or a desire to maximize their profit, Bangladeshi farmers are currently using pesticides in quantities that are 10-15 times higher than those recommended by the government. In general, Bangladeshi farmers do not follow the recommended dosages and apply pesticides in excessive amounts [<a href=\"#r-25\">25</a>]. Pesticide residues accumulate in the food chain, drinking water, and the environment as a result of this excessive application [<a href=\"#r-26\">26</a>]. It poses a significant health risk to present and future generations [<a href=\"#r-27\">27</a>]. In this study, a soil sample with a history (5-7 years) of shobicron and vertimec use was used to isolate a bacterial strain. We hypothesized that shobicron and vertimec would be digested and broken down by the bacteria.<br />\r\nThe toxicity of a pesticide is assessed by administering different doses of the active ingredient (a.i.) and each of its manufactured products to test animals. The active ingredient is the pesticide’s chemical component that controls the pest. The toxicities of different active components and different formulations containing the same active ingredient can be compared using LD<sub>50</sub> and LC<sub>50</sub> values. The lower the LD<sub>50</sub> or LC<sub>50</sub> of a pesticide, the more hazardous it is to people and animals. As a result, we first looked at the toxicity profiles of the pesticides on <em>Artemia salina.</em> After 36 hours of exopuse, the LC<sub>50</sub> values for Shobicron and vertimec were calculated to be 2.454323 g/ml and 2.210981 g/ml, respectively, and are considered extremely toxic to humans and animals.<br />\r\nWe used toxic shobicron and vertimec as a sole carbon source for the isolated bacteria to test detoxification efficiency. Research indicated that certain type of bacterial species have the capability to exploit sulfadoxine [<a href=\"#r-28\">28</a>], organophophorus [<a href=\"#r-29\">29</a>], tributyl phosphate [<a href=\"#r-30\">30</a>], chlorpyrifos [<a href=\"#r-31\">31</a>], and organophosphorous [<a href=\"#r-32\">32</a>] pesticides as carbon and energy source. In this study, morphological and biochemical tests were also carried out, and the isolated strain was gram negative, rod shaped, as previously identifying characteristics of <em>burkholderia </em>genus [<a href=\"#r-33\">33</a>]. <em>Burkholderia</em> is a highly versatile bacterium, which is capable of degradation of many hazardous pollutants [<a href=\"#r-34\">34, 35</a>], and it has a role in protection and growth promotion of plants [<a href=\"#r-36\">36, 37</a>].<br />\r\nIn this research, penicillin, kanamycin, gentamycin, tetracycline, ciprofloxacin, cefepime, carbapenem were used to test antibiotic sensitivity of the isolated bacteria. The result showed that strain was susceptible (S) to Cefepime and Penicillin but intermediate (I) resistant to Carbapenem, & Tetracycline and resistant (R) to Ciprofloxacin, Kanamycin, and Gentamycin. Previous research suggested that the strain resistant to gentamycin and ciprofloxacin antibiotics; and were sensitive to penicillin, cefepime [<a href=\"#r-38\">38, 39</a>].<br />\r\nBacteria that degrade pesticides have been found so far, and they comprise both Gram positive and Gram negative bacteria [<a href=\"#r-40\">40</a>]. Only a gram-negative bacterium was identified in this study. Atikur et al. found that two bacterial strains, Achromobacter and Diaphorobacter, can biodegrade pesticides [<a href=\"#r-41\">41</a>]. This was backed up by other scientists [<a href=\"#r-42\">42, 43</a>]. The majority of previously detected bacteria could only digest one pesticide. However, in this case, we identified one strain that could breakdown two pesticides. In its ideal state, this strain could breakdown both pesticides with an over 75% degradation rate. Similar results (Carbofuran degradation rate of about 80%) were obtained by Ekram et al. [<a href=\"#r-44\">44</a>]. As a result, this research has a lot of promise in terms of pesticide bioremediation. The current study was carried out in a laboratory setting. As a result, <em>Burkholderia Cepacia</em>-Like MB-01 should be evaluated with industrial waste and in natural habitats in future investigations. Furthermore, before being used to detoxify pesticides in the environment, <em>Burkholderia Cepacia</em>-Like MB-01 should be examined for pathogenicity to aquatic organisms like as fish, prawns, and crab [<a href=\"#r-45\">45, 46</a>].</p>"
},
{
"section_number": 5,
"section_title": "CONCLUSION",
"body": "<p>Environmental pollution is being brought on by Bangladesh’s massive rise in pesticide use. Reducing the amount of pesticide in the environment may be accomplished by bioremediation. The effectiveness of <em>Burkholderia Cepacia</em>-Like MB-01 in biodegrading shobicron and vertimec is revealed by this study, and it may be helpful for bioremediation programs aimed at enhancing soil quality. This study will enable many researchers to explore the crucial aspects of employing the natural isolate <em>Burkholderia Cepacia</em>-Like MB-01 for shobicron and vertimec biodegradation. As a result, a novel theory about the use of <em>Burkholderia Cepacia</em>-Like MB-01 to clean up pesticide-contaminated soil can develop.</p>"
},
{
"section_number": 6,
"section_title": "ACKNOWLEDGMENT",
"body": "<p>None</p>"
},
{
"section_number": 7,
"section_title": "AUTHOR CONTRIBUTIONS",
"body": "<p>MMZ, MMJ, MMM, SI, MJP, JB, AI: Methodology, Validation, Formal analysis, Investigation, Data curation, Writing–original draft. MSU, MAS: Conceptualization, Validation, Resources, Data curation, Project administration, Editing. SZ: Supervision, Funding acquisition.</p>"
},
{
"section_number": 8,
"section_title": "CONFLICTS OF INTEREST",
"body": "<p>There is no conflict of interest among the authors.</p>"
}
],
"figures": [
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/57/26/178-1652146042-Figure3.jpg",
"caption": "Figure 3. Optimization of pH and temperature. (a) Effect of pH on growth of the strain MB-01, where the maximum growth was obtained at pH 7; (b) Effect of temperature on bacterial growth, where maximum growth was observed at 35 °C. Data were recorded after 12 hours of incubation.",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/57/26/178-1652146042-Figure4.jpg",
"caption": "Figure 4. Antibiotic sensitivity test of the strain MB-01. Isolate was resistant to most of the tested antibiotics, and susceptible to Cefepime, and Penicillin. Note: Resistant=<10 mm; Intermediate resistance=10-15 mm; Susceptible=>15 mm.",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/57/26/178-1652146042-Figure5.jpg",
"caption": "Figure 5. Showing of degradation of shobicron (a) and vertimec (b) by the strain MB-01 in its optimum conditions. Data were recoded up to 5 days of incubation.",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/57/26/178-1652146042-Figure6.jpg",
"caption": "Figure 6. Effect of pH (a) and temperature (b) on shobicron and vertimec degradation by the strain MB-01. Data were recorded after 5 days of incubation.",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/57/26/178-1652146042-Figure1.jpg",
"caption": "Figure 1. Isolated shobicrn and vertimec degrading bacterial strain. (a) Bacterial growth on MS agar plate supplemented with 4 µg/ml of shobicron. (b) Gram staining isolated strain indicating gram-negative and rod-shaped structure.",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/57/26/178-1652146042-Figure2.jpg",
"caption": "Figure 2. Phylogenetic tree analysis with similar bacterial strains. The tree was constructed by the neighbor-joining method based on 16S rRNA gene sequences of strain MB-01 and related Burkholderia type strain.",
"featured": false
}
],
"authors": [
{
"id": 511,
"affiliation": [
{
"affiliation": "Microbiology Laboratory, Department of Genetic Engineering and Biotechnology, University of Rajshahi, Rajshahi- 6205, Bangladesh"
}
],
"first_name": "Md.",
"family_name": "Moniruzzaman",
"email": null,
"author_order": 1,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 127
},
{
"id": 512,
"affiliation": [
{
"affiliation": "Department of Mathematics and Natural Sciences, Brac University, Dhaka- 1212, Bangladesh"
}
],
"first_name": "Mst. Maskera",
"family_name": "Jinnah",
"email": null,
"author_order": 2,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 127
},
{
"id": 513,
"affiliation": [
{
"affiliation": "Microbiology Laboratory, Department of Genetic Engineering and Biotechnology, University of Rajshahi, Rajshahi- 6205, Bangladesh"
}
],
"first_name": "Most. Merina",
"family_name": "Mostari",
"email": null,
"author_order": 3,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 127
},
{
"id": 514,
"affiliation": [
{
"affiliation": "Microbiology Laboratory, Department of Genetic Engineering and Biotechnology, University of Rajshahi, Rajshahi- 6205, Bangladesh"
}
],
"first_name": "Shirmin",
"family_name": "Islam",
"email": null,
"author_order": 4,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 127
},
{
"id": 515,
"affiliation": [
{
"affiliation": "Microbiology Laboratory, Department of Genetic Engineering and Biotechnology, University of Rajshahi, Rajshahi- 6205, Bangladesh"
}
],
"first_name": "Md. Joy",
"family_name": "Pramanik",
"email": null,
"author_order": 5,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 127
},
{
"id": 516,
"affiliation": [
{
"affiliation": "Microbiology Laboratory, Department of Genetic Engineering and Biotechnology, University of Rajshahi, Rajshahi- 6205, Bangladesh"
}
],
"first_name": "Jui",
"family_name": "Biswas",
"email": null,
"author_order": 6,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 127
},
{
"id": 517,
"affiliation": [
{
"affiliation": "Microbiology Laboratory, Department of Genetic Engineering and Biotechnology, University of Rajshahi, Rajshahi- 6205, Bangladesh"
}
],
"first_name": "Al",
"family_name": "Imran",
"email": null,
"author_order": 7,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 127
},
{
"id": 518,
"affiliation": [
{
"affiliation": "Microbiology Laboratory, Department of Genetic Engineering and Biotechnology, University of Rajshahi, Rajshahi- 6205, Bangladesh"
}
],
"first_name": "Md. Salah",
"family_name": "Uddin",
"email": null,
"author_order": 8,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 127
},
{
"id": 519,
"affiliation": [
{
"affiliation": "Microbiology Laboratory, Department of Genetic Engineering and Biotechnology, University of Rajshahi, Rajshahi- 6205, Bangladesh"
}
],
"first_name": "Md. Abu",
"family_name": "Saleh",
"email": "saleh@ru.ac.bd",
"author_order": 9,
"ORCID": null,
"corresponding": true,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "Md. Abu Saleh, PhD; Microbiology Laboratory, Department of Genetic Engineering and Biotechnology, University of Rajshahi, Rajshahi- 6205, Bangladesh, e-mail: saleh@ru.ac.bd",
"article": 127
},
{
"id": 520,
"affiliation": [
{
"affiliation": "Microbiology Laboratory, Department of Genetic Engineering and Biotechnology, University of Rajshahi, Rajshahi- 6205, Bangladesh"
}
],
"first_name": "Shahriar",
"family_name": "Zaman",
"email": "szaman@ru.ac.bd",
"author_order": 10,
"ORCID": null,
"corresponding": true,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "Shahriar Zaman, Microbiology Laboratory, Department of Genetic Engineering and Biotechnology, University of Rajshahi, Rajshahi- 6205, Bangladesh,\r\ne-mail: szaman@ru.ac.bd",
"article": 127
}
],
"views": 1098,
"downloads": 106,
"references": [
{
"id": 4185,
"serial_number": 1,
"pmc": null,
"reference": "Glazer AN, Nikaido H. Microbial biotechnology: fundamentals of applied microbiology. Cambridge University Press; 2007.",
"DOI": null,
"article": 127
},
{
"id": 4186,
"serial_number": 2,
"pmc": null,
"reference": "Rani K, Dhania G. Bioremediation and biodegradation of pesticide from contaminated soil and water—a noval approach. Int J Curr Microbiol App Sci. 2014; 3(10): 23-33.",
"DOI": null,
"article": 127
},
{
"id": 4187,
"serial_number": 3,
"pmc": null,
"reference": "Zaller JG. Daily Poison: Pesticides-an Underestimated Danger. Springer Nature; 2020.",
"DOI": null,
"article": 127
},
{
"id": 4188,
"serial_number": 4,
"pmc": null,
"reference": "Singh B. Role of pesticides in management of crop pests. Jodhpur: Scientific Publishers (India); 2012.",
"DOI": null,
"article": 127
},
{
"id": 4189,
"serial_number": 5,
"pmc": null,
"reference": "Alam MK. Determination of cypermethrin, chlorpyrifos and diazinon residues in tomato and reduction of cypermethrin residues in tomato using rice bran. World. 2013; 1(2): 30-5.",
"DOI": null,
"article": 127
},
{
"id": 4190,
"serial_number": 6,
"pmc": null,
"reference": "World Health Organization. Preventing disease through healthy environments: exposure to lead: a major public health concern. World Health Organization. License: CC BY-NC-SA 3.0 IGO. 2019",
"DOI": null,
"article": 127
},
{
"id": 4191,
"serial_number": 7,
"pmc": null,
"reference": "Khan WA, Ali S, Shah SA. Water Pollution: Sources and Its Impact on Human Health, Control and Managing. Journal of International Cooperation and Development. 2022;5(1): 69-69.",
"DOI": null,
"article": 127
},
{
"id": 4192,
"serial_number": 8,
"pmc": null,
"reference": "Patel P, Prajapati R. Possible Toxicological Effect of Pesticides, Cypermethrin on Earthworm (Esiania fetida). International Journal of Research in Engineering, Science and Management. 2020; 3(12): 138-43.",
"DOI": null,
"article": 127
},
{
"id": 4193,
"serial_number": 9,
"pmc": null,
"reference": "Korkmaz V, Güngördü A, Ozmen M. Comparative evaluation of toxicological effects and recovery patterns in zebrafish (Danio rerio) after exposure to phosalone-based and cypermethrin-based pesticides. Ecotoxicology and Environmental Safety. 2018; 160: 265-72.",
"DOI": null,
"article": 127
},
{
"id": 4194,
"serial_number": 10,
"pmc": null,
"reference": "Singh RP, Singh R. Adsorption and movement of cypermethrin on Indian soils amended with cationic, non-ionic and anionic surfactants. Adsorption Science & Technology. 2004; 22(7): 553-64.",
"DOI": null,
"article": 127
},
{
"id": 4195,
"serial_number": 11,
"pmc": null,
"reference": "Al-Autaish HHN. Clinical study on toxicity of cypermethrin in Arrabi sheep. Doctoral dissertation, M. Sc. Thesis, Coll. of Veterinary Medicine, Univ. of Basrah. 2010; pp: 76.",
"DOI": null,
"article": 127
},
{
"id": 4196,
"serial_number": 12,
"pmc": null,
"reference": "Guo J, Tong M, Tang J, Bian H, Wan X, He L, et al. Analysis of multiple pesticide residues in polyphenol-rich agricultural products by UPLC-MS/MS using a modified QuEChERS extraction and dilution method. Food chemistry. 2019; 274: 452-9.",
"DOI": null,
"article": 127
},
{
"id": 4197,
"serial_number": 13,
"pmc": null,
"reference": "Ali SW, Li R, Zhou WY, Sun JQ, Guo P, Ma JP, et al. Isolation and characterization of an abamectin-degrading Burkholderia cepacia-like GB-01 strain. Biodegradation. 2010; 21(3): 441-52.",
"DOI": null,
"article": 127
},
{
"id": 4198,
"serial_number": 14,
"pmc": null,
"reference": "Suryawanshi V, Yadav A, Mohite S, Magdum CS. Toxicological Assessment using Brine Shrimp Lethality Assay and Antimicrobial activity of Capparis Grandis. Journal of University of Shanghai for Science and Technology. 2020; 22(11): 746-59.",
"DOI": null,
"article": 127
},
{
"id": 4199,
"serial_number": 15,
"pmc": null,
"reference": "Lyne PM, Grange JM. Collins and Lyne’s microbiological methods. 8th Edition, Oxford University Press, New York. 2004.",
"DOI": null,
"article": 127
},
{
"id": 4200,
"serial_number": 16,
"pmc": null,
"reference": "Bauer AW. Antibiotic susceptibility testing by a standardized single disc method. Am J clin pathol. 1966; 45: 149-58.",
"DOI": null,
"article": 127
},
{
"id": 4201,
"serial_number": 17,
"pmc": null,
"reference": "Bwapwa JK. Factors affecting the bioremediation of industrial and domestic wastewaters. In Microbial Biodegradation and Bioremediation, Elsevier. 2022; (pp. 461-472).",
"DOI": null,
"article": 127
},
{
"id": 4202,
"serial_number": 18,
"pmc": null,
"reference": "Singh R. Microorganism as a tool of bioremediation technology for cleaning environment: a review. Proceedings of the International Academy of Ecology and Environmental Sciences. 2014; 4(1): 1.",
"DOI": null,
"article": 127
},
{
"id": 4203,
"serial_number": 19,
"pmc": null,
"reference": "Harms H, Schlosser D, Wick LY. Untapped potential: exploiting fungi in bioremediation of hazardous chemicals. Nature Reviews Microbiology. 2011; 9(3): 177-92.",
"DOI": null,
"article": 127
},
{
"id": 4204,
"serial_number": 20,
"pmc": null,
"reference": "Hossain K, Ismail N. Bioremediation and detoxification of pulp and paper mill effluent: A review. Research Journal of Environmental Toxicology. 2015; 9(3): 113.",
"DOI": null,
"article": 127
},
{
"id": 4205,
"serial_number": 21,
"pmc": null,
"reference": "Singh BK, Walker A, Wright DJ. Bioremedial potential of fenamiphos and chlorpyrifos degrading isolates: influence of different environmental conditions. Soil Biology and Biochemistry. 2006; 38(9): 2682-93.",
"DOI": null,
"article": 127
},
{
"id": 4206,
"serial_number": 22,
"pmc": null,
"reference": "Md Z, Mohammed A, Badrul I, Azza SA, Mohamed AE, Arif A. Bioremediation: A tool for environmental cleaning. African Journal of Microbiology Research. 2009; 3(6): 310-4.",
"DOI": null,
"article": 127
},
{
"id": 4207,
"serial_number": 23,
"pmc": null,
"reference": "Huang Y, Xiao L, Li F, Xiao M, Lin D, Long X, et al. Microbial degradation of pesticide residues and an emphasis on the degradation of cypermethrin and 3-phenoxy benzoic acid: a review. Molecules. 2018; 23(9): 2313.",
"DOI": null,
"article": 127
},
{
"id": 4208,
"serial_number": 24,
"pmc": null,
"reference": "Hussain A, Rehman F, Rafeeq H, Waqas M, Asghar A, Afsheen N, et al. In-situ, Ex-situ, and nano-remediation strategies to treat polluted soil, water, and air–A review. Chemosphere. 2022; 289: 133252.",
"DOI": null,
"article": 127
},
{
"id": 4209,
"serial_number": 25,
"pmc": null,
"reference": "Chowdhury MG, Rahman MA, Miaruddin M, Khan MH, Rahman MM. Assessment of pesticides and ripening chemicals used in selected vegetables at different locations of Bangladesh. Bangladesh Journal of Agricultural Research. 2019; 44(2): 261-79.",
"DOI": null,
"article": 127
},
{
"id": 4210,
"serial_number": 26,
"pmc": null,
"reference": "Liu L, Zheng X, Wei X, Kai Z, Xu Y. Excessive application of chemical fertilizer and organophosphorus pesticides induced total phosphorus loss from planting causing surface water eutrophication. Scientific Reports. 2021; 11(1): 1-8.",
"DOI": null,
"article": 127
},
{
"id": 4211,
"serial_number": 27,
"pmc": null,
"reference": "Rahman S. Farm-level pesticide use in Bangladesh: determinants and awareness. Agriculture, ecosystems & environment. 2003; 95(1): 241-52.",
"DOI": null,
"article": 127
},
{
"id": 4212,
"serial_number": 28,
"pmc": null,
"reference": "Zhang W, Xu D, Niu Z, Yin K, Liu P, Chen L. Isolation and characterization of Pseudomonas sp. DX7 capable of degrading sulfadoxine. Biodegradation. 2012; 23(3): 431-9.",
"DOI": null,
"article": 127
},
{
"id": 4213,
"serial_number": 29,
"pmc": null,
"reference": "Chino-Flores C, Dantán-González E, Vázquez-Ramos A, Tinoco-Valencia R, Díaz-Méndez R, Sánchez-Salinas E, et al. Isolation of the opdE gene that encodes for a new hydrolase of Enterobacter sp. capable of degrading organophosphorus pesticides. Biodegradation. 2012; 23(3): 387-97.",
"DOI": null,
"article": 127
},
{
"id": 4214,
"serial_number": 30,
"pmc": null,
"reference": "Ahire KC, Kapadnis BP, Kulkarni GJ, Shouche YS, Deopurkar RL. Biodegradation of tributyl phosphate by novel bacteria isolated from enrichment cultures. Biodegradation. 2012; 23(1): 165-76.",
"DOI": null,
"article": 127
},
{
"id": 4215,
"serial_number": 31,
"pmc": null,
"reference": "Harishankar MK, Sasikala C, Ramya M. Efficiency of the intestinal bacteria in the degradation of the toxic pesticide, chlorpyrifos. 3. Biotech. 2013; 3(2): 137-42.",
"DOI": null,
"article": 127
},
{
"id": 4216,
"serial_number": 32,
"pmc": null,
"reference": "Kanekar PP, Bhadbhade BJ, Deshpande NM, Sarnaik SS. Biodegradation of organophosphorus pesticides. Proceedings-Indian National Science Academy Part B. 2004; 70(1): 57-70.",
"DOI": null,
"article": 127
},
{
"id": 4217,
"serial_number": 33,
"pmc": null,
"reference": "AuCoin DP, Crump RB, Thorkildson P, Nuti DE, LiPuma JJ, Kozel TR. Identification of Burkholderia cepacia complex bacteria with a lipopolysaccharide-specific monoclonal antibody. Journal of medical microbiology. 2010; 59(Pt 1): 41.",
"DOI": null,
"article": 127
},
{
"id": 4218,
"serial_number": 34,
"pmc": null,
"reference": "Morya R, Salvachúa D, Thakur IS. Burkholderia: an untapped but promising bacterial genus for the conversion of aromatic compounds. Trends in Biotechnology. 2020; 38(9): 963-75.",
"DOI": null,
"article": 127
},
{
"id": 4219,
"serial_number": 35,
"pmc": null,
"reference": "Seo JS, Keum YS, Li QX. Bacterial degradation of aromatic compounds. International journal of environmental research and public health. 2009; 6(1): 278-309.",
"DOI": null,
"article": 127
},
{
"id": 4233,
"serial_number": 36,
"pmc": null,
"reference": "Zaman NR, Chowdhury UF, Reza RN, Chowdhury FT, Sarker M, Hossain MM, et al. Plant growth promoting endophyte Burkholderia contaminans NZ antagonizes phytopathogen Macrophomina phaseolina through melanin synthesis and pyrrolnitrin inhibition. PloS one. 2021; 16(9): e0257863.",
"DOI": null,
"article": 127
},
{
"id": 4234,
"serial_number": 37,
"pmc": null,
"reference": "Kong P, Hong C. Endophytic Burkholderia sp. SSG as a potential biofertilizer promoting boxwood growth. PeerJ. 2020; 8: e9547.",
"DOI": null,
"article": 127
},
{
"id": 4235,
"serial_number": 38,
"pmc": null,
"reference": "Roy P, Ahmed N, Biswal I, Grover RK. Antimicrobial susceptibility pattern of Burkholderia cepacia isolates from patients with malignancy. Journal of Global Infectious Diseases. 2014; 6(2): 90.",
"DOI": null,
"article": 127
},
{
"id": 4236,
"serial_number": 39,
"pmc": null,
"reference": "Moore JE, Crowe M, Shaw A, McCaughan J, Redmond AO, Elborn JS. Antibiotic resistance in Burkholderia cepacia at two regional cystic fibrosis centres in Northern Ireland: is there a need for synergy testing?. Journal of Antimicrobial Chemotherapy. 2001; 48(2): 319-21.",
"DOI": null,
"article": 127
},
{
"id": 4237,
"serial_number": 40,
"pmc": null,
"reference": "DeLorenzo ME, Scott GI, Ross PE. Toxicity of pesticides to aquatic microorganisms: a review. Environmental Toxicology and Chemistry: An International Journal. 2001; 20(1): 84-98.",
"DOI": null,
"article": 127
},
{
"id": 4238,
"serial_number": 41,
"pmc": null,
"reference": "Rahman MA, Arefin AS, Saha O, Rahaman MM. Isolation and identification of pesticides degrading bacteria from farmland soil. Bangladesh Journal of Microbiology. 2018; 35(2): 90-4.",
"DOI": null,
"article": 127
},
{
"id": 4239,
"serial_number": 42,
"pmc": null,
"reference": "Ren HF, Li SQ, Liu SJ, Liu ZP. Isolation and characterization of a p-chloroaniline-degrading bacterial strain. Huan Jing ke Xue= Huanjing Kexue. 2005; 26(1): 154-8.",
"DOI": null,
"article": 127
},
{
"id": 4240,
"serial_number": 43,
"pmc": null,
"reference": "Xia ZY, Zhang L, Zhao Y, Yan X, Li SP, Gu T, et al. Biodegradation of the herbicide 2, 4-dichlorophenoxyacetic acid by a new isolated strain of Achromobacter sp. LZ35. Current microbiology. 2017; 74(2): 193-202.",
"DOI": null,
"article": 127
},
{
"id": 4241,
"serial_number": 44,
"pmc": null,
"reference": "Ekram MA, Sarker I, Rahi MS, Rahman MA, Saha AK, Reza MA. Efficacy of soil‐borne Enterobacter sp. for carbofuran degradation: HPLC quantitation of degradation rate. Journal of basic microbiology. 2020; 60(5): 390-9.",
"DOI": null,
"article": 127
},
{
"id": 4242,
"serial_number": 45,
"pmc": null,
"reference": "Ortiz-Hernández ML, Sánchez-Salinas E, Dantán-González E, Castrejón-Godínez ML. Pesticide biodegradation: mechanisms, genetics and strategies to enhance the process. Biodegradation-life of Science. 2013; 10: 251-87.",
"DOI": null,
"article": 127
},
{
"id": 4243,
"serial_number": 46,
"pmc": null,
"reference": "Ravi RK, Bhawana P, Fulekar MH. Bioremediation of persistent pesticides in rice field soil environment using surface soil treatment reactor. International Journal of Current Microbiology and Applied Sciences. 2015; 4(2): 359-69.",
"DOI": null,
"article": 127
}
]
},
{
"id": 129,
"slug": "178-1650449385-molecular-characterization-and-antibacterial-activities-of-mangrove-endophytic-fungi-from-coastal-kenya",
"featured": false,
"slider": false,
"issue": "Vol5 Issue3",
"type": "original_article",
"manuscript_id": "178-1650449385",
"recieved": "2022-04-21",
"revised": null,
"accepted": "2022-07-07",
"published": "2022-07-26",
"pdf_file": "https://jabet.bsmiab.org/media/pdf_file/2023/33/178-1650449385.pdf",
"title": "Molecular characterization and antibacterial activities of mangrove endophytic fungi from coastal Kenya",
"abstract": "<p>The advent of resistant pathogenic bacteria and fungi across the globe is threatening the efficacy of antibiotic drugs. Thus, microbial infections are becoming a threat to life. Endophytic fungi remain a viable source of secondary metabolites with unique spectra of biological activities. This study isolated and characterized endophytic fungi from selected mangrove species of coastal Kenya and further ascertained their activities. A total of 18 fungal endophytes selected from mangrove species were investigated for antimicrobial activity against gram-positive <em>Staphylococcus aureus </em>and gram-negative <em>Escherichia coli</em>. Potato dextrose agar and potato dextrose broth were used for isolation, purification, and fermentation at 28<sup>o</sup>C for 7–15 days. Extraction of fungal metabolites was achieved using ethyl acetate (1:1 v/v) and ethyl acetate in 10% methanol (9:1 v/v). Solvents were recovered in a fume hood and extracts were dissolved in 1 ml of dimethyl sulfoxide. Molecular characterization completely identified 9 species, namely: <em>Aspergillus</em> <em>flavus, Aspergillus niger, Aspergillus tubingensis, Aspergillus oryzae, Rhizophora nomius, Aspergillus awamori, Aspergillus aculeatus, Aspergillus bravionivious, and Aspergillus welwitchiae. </em>The minimum inhibitory concentration of ethyl acetate crude extracts of the most active fungal isolate, <em>A. flavus </em>(MT447532.1), was 0.91 ± 0.05 mg/ml and 0.82 ± 0.052 mg/ml against <em>S. aureus </em>and <em>E. coli</em>, respectively. Results showed that some crude extracts of mangrove fungal endophytes from coastal Kenya are effective against bacteria, hence a promising source of novel organic natural metabolites with a possible wide range of biological activities.</p>",
"journal_reference": "J Adv Biotechnol Exp Ther. 2022; 5(3): 654-666.",
"academic_editor": "Akhi Moni, PhD; ABEx Bio-Research Center, Dhaka-1230, Bangladesh",
"cite_info": "Kiti HM, Kibiti CM, et al. Molecular characterization and antibacterial activities of mangrove endophytic fungi from coastal Kenya. J Adv Biotechnol Exp Ther. 2022; 5(3): 654-666.",
"keywords": [
"Antibacterial activity",
"Endophytic fungi",
"Molecular characterization",
"Kenya coast"
],
"DOI": "doi.org/10.5455/jabet.2022.d144",
"sections": [
{
"section_number": 1,
"section_title": "INTRODUCTION",
"body": "<p>There has been a rapid increase in multidrug-resistant (MDR) bacteria against antibiotics. This being a worldwide phenomenon, new resistance mechanisms are emerging and a rise in MDR is one of the reasons for the failure of treatment and increased deaths [<a href=\"#r-1\">1</a>]. The emergence and spread of drug-resistant pathogens continue to threaten the treatment of common infections. The World Health Organization’s (WHO) reserves of novel antimicrobials are being depleted, making access to quality medicines a challenge [<a href=\"#r-2\">2</a>]. To date, pathogenic strains of bacteria that were previously treatable through antibiotic therapy are resistant to every available antibiotic, while many other serious and pathogenic strains are resistant to all but one drug, thereby placing them on the edge of being deadly pathogens [<a href=\"#r-3\">3</a>]. <em>Staphylococcus aureus </em>is an adaptable pathogen found in both animals and human beings and is responsible for increasing infections, starting with minor skin infections, and progressing to more serious and deadly ones such as meningitis and pneumonia. This pathogen is an important agent of food poisoning globally and is transmitted to humans through contaminated and unprocessed milk and milk products. It is resistant to penicillin, methicillin, and vancomycin [<a href=\"#r-4\">4, 5</a>]. <em>Escherichia coli</em> is a rodlike gram-negative bacterium commonly occupying the lower intestine of mammals and whose transmission is mostly by fecal-oral-rout [6]. Most <em>E. coli</em> are innocuous normal flora in the gut that are beneficial to their host through the production of Vitamin K<sub>2 </sub>and B-Complex. These vitamins aid in preventing the establishment of bacteria within the intestinal walls. However, others, such as serotype O157:H7, cause food poisoning in humans, hemolytic uremic syndrome (HUS), and death in the elderly, the very young, and immunocompromised individuals. Antibiotic resistance is on the rise because of antibiotic overuse in humans and the rate of adaptive mutations in <em>E. coli</em>, an important factor for bacterial antibiotic resistance [<a href=\"#r-6\">6</a>].<br />\r\nNatural products have been the utmost source of all medicines for most of human history [<a href=\"#r-7\">7</a>]. Endophytic fungi inhabit an unmatched environmental position, being thought to impact the ecology, distribution of plants, physiology, and biochemistry [<a href=\"#r-7\">7</a>]. Fungal endophytes have also been known as possible sources of innovative naturally occurring compounds applicable in agricultural, industrial, and pharmaceutical applications, especially those secondary metabolites produced by fungal endophytes inhabiting medicinal plants [<a href=\"#r-8\">8</a>]. Mangrove endophytic fungi have become an important natural source for the discovery of novel biologically active compounds [<a href=\"#r-9\">9</a>]. Mangroves are uniquely adapted to harsh environmental conditions like high salinity, anaerobic soils, and the rising sea levels [<a href=\"#r-10\">10</a>]. They function as primary producers in the estuarine food chain, contributing to an ecological, economic, scientific, and cultural resource [<a href=\"#r-10\">10</a>]. Most mangrove genera and families are not closely related but have common adaptive features. These include developed morphological, physiological, biological, and ecological adaptations to extreme environmental conditions such as changes in sea level, high salinity, high temperatures, and anaerobic soils [<a href=\"http://v1\">11</a>]. This is made possible by the presence of such features including pneumatophores, stilt roots, salt-secreting leaves, and viviparous water-dispersed propagules [<a href=\"#r-10\">10</a>]. In Kenya, there are approximately 54,000 ha of mangrove forests, most of which are found in north coast in the counties of Tana River and Lamu [<a href=\"#r-12\">12</a>]. Endophytic fungi inhabit the internal tissues of host plants for all or part of their life cycle. Almost every plant species harbors non-mycorrhizal fungal endophytes [<a href=\"#r-8\">8</a>]. Among the diverse species of mangroves, only 9 are reported to occur along the Kenya coast, where their ecological habitats are systematically being destroyed through human activities [<a href=\"#r-13\">13</a>], thus threatening them with imminent extinction. The diversity of mangrove fungal endophytes of <em>Rhizophora mucronata</em>, <em>Avicenia marina</em>, <em>Ceriops tagal, </em>and <em>Sonneratia alba</em> were investigated based on cultural and morphological characteristics [<a href=\"#r-14\">14</a>]. A total of 18 fungal isolates based on the cultural and morphological characterization have been reported [<a href=\"#r-14\">14</a>].<br />\r\nThe present study focused on molecular characterization of the mangrove endophytic fungi and their antibacterial potential of crude extracts from selected mangrove species.</p>"
},
{
"section_number": 2,
"section_title": "MATERIALS AND METHODS",
"body": "<p><strong>Description of the study area</strong><br />\r\nThe study was conducted along the Kenya coast in three different mangrove forests (<a href=\"#figure1\">Figure 1</a>). Gazi Bay is located on the south coast of Kenya, at 4°25’S and 39°50’E in Kwale County, about 50 km south of Mombasa. This bay is made up of a shallow channel measuring about 4 km long, bordered by mangrove forests dominated by the species <em>Sonneratia alba </em>and<em> Rhizophora mucronata </em>[<a href=\"#r-15\">15</a>]. Tudor Creek is situated at 42’S, 3940’E on northwest of Mombasa Island and extends between 10–15 km inland. Two seasonal rivers, Kombeni and Tsalu, drain into the creek over 45,000 and 10,000 ha, respectively [<a href=\"#r-16\">16</a>]. A mangrove forest within the creek extends over an area of 1, 1.64 ha, primarily made up of <em>Sonneratia alba</em>, <em>Avicenia</em> <em>marina, </em>and<em> Rhizophora mucronata</em>. The forest has different mangrove species zonation along the tidal gradient and is covered by deposits that are mostly made up of mud and sand. Parts of the forest are made up of mud and sand [<a href=\"#r-17\">17</a>]. Mida Creek (03°21’S, 39°59’E) is situated in Kilifi County approximately 25 km south of Malindi town and about 88 km north of Mombasa City [<a href=\"#r-16\">16</a>]. The mangrove forest is the most prominent habitat in Mida Creek and occupies approximately 1,746 ha. The forest supports 7 out of the 9 mangrove species found in Kenya [<a href=\"#r-13\">13</a>].</p>\r\n\r\n<div id=\"figure1\">\r\n<figure class=\"image\"><img alt=\"\" height=\"715\" src=\"/media/article_images/2023/03/25/178-1650449385-Figure1.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 1.</strong> A map of the Kenya coast showing mangrove forests of Gazi Bay, Tudor and Mida creeks from where mangrove twig samples were collected [<a href=\"#r-14\">14</a>].</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Isolation of mangrove endophytic fungi</strong><br />\r\nStem twigs from selected mangrove species of <em>Sonneratia alba</em>, <em>Ceriops tagal</em>, <em>Rhizophora mucronata, </em>and <em>Avicenia marina</em> were collected and transported in airtight bags to the laboratory and stored. Surface sterilization was achieved by washing them in running tap water to remove debris. They were dipped in 70% for 3 min and then in 4% sodium hypochlorite for 3 min, followed by rinsing with sterile water for 2 min and drying under sterile conditions [<a href=\"#r-14\">14</a>]. The outer tissues were removed using a laboratory knife after drying the plant tissues under sterile laminar air flow and passing through a flame. Internal tissues were split into smaller pieces of 0.5 to 1 cm under sterile conditions and plated on Potato Dextrose Agar (PDA) in order to obtain a wide range of fungal isolates. The plates were then incubated for 7-10 days at 28<sup>o</sup>C. Fungal growth was observed and monitored for the 10 days. Purity plating was done to get pure colonies, and it was done in triplicates for molecular characterization [<a href=\"#r-14\">14</a>].</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Molecular identification of the mangrove fungal endophytes</strong><br />\r\nMangrove fungal endophytes were selected for molecular characterization based on their growth characteristics such as color, colony form, mycelium, and colony margin. A total of 18 mangrove endophytic fungal isolates from the selected mangrove species of <em>Sonneratia alba, Ceriops tagal, Rhizophora mucronata, </em>and <em>Avicenia marina </em>were chosen for molecular identification. Total genomic deoxyribonucleic acid (DNA) extraction of selected mangrove fungal endophytes was done using the <em>Quick-</em>DNA<sup>TM</sup> Fungal/Bacterial Mini Preparation Kit by Zymo Research, Catalog No. D6005 [<a href=\"#r-18\">18</a>]. Prior to that, selected mangrove fungal endophytes were cultured in PDA at 28<sup>o</sup>C for 5 days. To remove any traces of DNA, all surfaces were cleaned with 3.85 % sodium hypochlorite, followed by 70% ethanol. The extraction buffers were carefully prepared using deionized water and later sterilized by autoclaving at 121<sup>o</sup>C for 15 minutes. All reusable apparatus was first UV-sterilized for 1 hour before being cleaned in between by soaking in 3.85% sodium hypochlorite for 30 minutes and thoroughly rinsing with sterilized distilled water. Beta-mercaptoethanol was added to the Genomic Lysis Buffer at a final concentration of 0.5% (v/v) to improve performance [<a href=\"#r-18\">18</a>].<br />\r\nThe target regions were amplified using universal primers for fungal DNA; internal transcribed spacers, ITS1 F-(5″-CTTGGTCATTTAGAGGAAGTAA-3″) and ITS4 (5″-TCCTCCGCTTATTGATATGC-3″). Agarose gel was prepared according to standard procedures and then mounted on the gel electrophoresis tank. 3 µl of the sample was mixed with a drop of loading dye and loaded into the wells, and electric current was run at 70 V for 50 minutes. The gel was then observed under a UV-Transilluminator, and gel images were captured. The concentration and purity of the DNA were confirmed using spectrophotometry (Eppendorf Bio spectrometer, software 4.3.5.0). From the agarose gel electrophoresis, DNA bands of sufficient thickness were observed, a preliminary indicator of a successful DNA extraction. All working areas were cleaned with 3.85% sodium hypochlorite to remove any traces of DNA, followed by 70% methanol for quality control. The polymerase chain reaction (PCR) amplicons were sequenced at Inqaba Boitech (South Africa). The DNA sequences obtained from ITS amplification were submitted to GenBank, and accession numbers were assigned to each fungus. DNA sequences were edited using UGENE version 36.0 and molecular evolutionary genetics analysis software for microorganisms (MEGA X).</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Fermentation and extraction of fungal metabolites</strong><br />\r\nAll endophytic cultures were cultivated in 250 ml universal bottles containing 30 ml of Potato Dextrose Broth (PDB) and incubated at 28<sup>o</sup>C for about 3 weeks under static conditions. The broth culture was then filtered to separate the mycelia from the filtrate. The cell-free fermentation broths were extracted using two methods. In one method, the broth was homogenized in a 10% methanol/water mixture before being extracted in analytical grade (98% purity) ethyl acetate. The other method involved homogenization of the fungal broth and extraction with ethyl acetate. In each case, the fungal mycelia were separated from the filtrate by filtering them through four layers of muslin clothing. The cell-free fermentation broths were extracted three times with equal volumes of ethyl acetate. The mixture was left to stand for two hours and filtered to remove the mycelium residues. The filtrate was collected in a 50 ml centrifuge tube and centrifuged at 8,000 revolutions per minute (rpm) for 10 minutes to separate the organic phase and the broth phase [<a href=\"#r-14\">14</a>]. The organic layer was removed by decantation and transferred to new tubes. An equal volume of ethyl acetate was added to the broth phase and extracted three more times. The resulting extracts were evaporated to dryness in a fume hood and gave a solid and oily residue. Extracts were dissolved in dimethyl sulfide oxide (DMSO) and used for antimicrobial assays. The DMSO extracts were kept at -20 °C for future research [<a href=\"#r-10\">10</a>].</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong><em>In vitro</em></strong><strong> antibacterial susceptibility test</strong><br />\r\nEthyl acetate and methanolic crude extracts of the mangrove fungal endophytes dried in the fume hood were tested against <em>S. aureus, </em>American Type Culture Collection (ATCC 25922) and <em>E.</em> coli (ATCC 25922) obtained from Technical University of Mombasa, (TUM) Biology laboratory. Antimicrobial activities of the mangrove endophytic fungal crude extracts were determined by agar disk diffusion method. The test organisms were prepared in Mueller Hinton Agar Nutrient broth and incubated at 37 ºC for 24 hours and 3 days at 28 ± 1ºC, respectively [<a href=\"#r-10\">10</a>]. Mueller Hinton Agar was prepared according to the manufacturer’s instructions. 32g of the Mueller Hinton agar was weighed and dissolved in 900 ml of distilled water. The mixture was heated to completely dissolve the medium, autoclaved at 121°C for 15 minutes and allowed to cool for 30 minutes. The media was poured on sterile petri dishes to 24 mm in a laminar flow hood to avoid any contamination and allowed to cool for a complete setting for 30 minutes. The plates were left ajar to avoid moisture build-up on the lid that could cause contamination. The plates were then appropriately labelled. After incubation, bacterial suspensions were spread on the surface of the solid agar medium (MHA) using sterile inoculating loop and left to dry for 5 minutes before being used for the antimicrobial assay [<a href=\"#r-10\">10</a>]. The discs were placed on the dried solid agar medium using a sterile pair of forceps and firmly pressed on the surface to prevent falling off during incubation. Discs were evenly placed on the surface of the medium ensuring that a distance of 12-15 mm away was maintained from the edge of the plate and away from each other. Each petri plate contained one test sample, one positive control Amoxicillin and one negative control (DMSO). Lids were replaced, sealed with Parafilm and plates were incubated at 37ºC for 24 hours and activity was expressed as the diameter of the clear zone of inhibition (mm) around agar plugs indicating antimicrobial activity of the crude mangrove endophytic fungal extracts [<a href=\"#r-19\">19, 20</a>].</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Minimum inhibition concentrations of </strong><strong><em>Aspergillus flavus</em></strong><strong> extracts</strong><br />\r\nMinimum inhibition concentrations (MICs) of the ethyl acetate crude extracts of <em>Aspergillus flavus</em> which displayed the strongest antimicrobial activity against S. aureus (ATCC 25922) and <em>E. </em>coli (ATCC 25922) were determined using micro-dilution method in a 96-well plate. The crude extract of the fungal isolate <em>A. flavus</em> was dissolved in DMSO and working concentrations made in 2-fold serial dilutions where the highest concentration was 1 mg/ml. The bacterial test organisms were grown on Müeller Hinton broth to 0.5 Macfarland standard. Test antibiotic solution was prepared as 1 g/ml then diluted in Müeller Hinton broth to 2-fold the top concentration desired in the test. 100 µL of Müeller Hinton broth was dispensed into all wells of the microtiter plate. 100 µL of 2-fold antibiotic solution was pipetted into the wells of column 1. Using the pipette, the antibiotic was mixed by sucking up and down 5-8 times. 100 µL of the mixture was withdrawn from column 1 and added to column 2 and then to column 3. The procedure was repeated up to column 10. 100 µL was discarded from column 10. 5 µL of bacteria was poured into wells from columns 1 to 11. No bacteria were added into column 12 serving as the blank. Plates were incubated at 37 °C for 18 hours. The reading of results was done in dual-mode microplate reader with monochromator-based optics for absorbance and sensitive top and bottom reading fluorescence applications (INFINITE M NANO) Pro200 Series, Austria at 600 nm. Experiments were performed in triplicate [<a href=\"#r-21\">21</a>].</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Data analysis</strong><br />\r\n<em>Phylogenetic analysis</em><br />\r\nThe DNA sequences with assigned accession numbers and sequence matches from National Centre for Biological Information (NCBI) Basic Local Alignment Search Tool (BLAST) were retrieved and combined into a single text file in fasta format. Using UGENE, NCBI BLAST, the data base was queried using default settings and percentage identity threshold set at about 97% [<a href=\"#r-22\">22</a>]. The sequences were aligned in Multi Sequence Alignment (MUSCLE) software found in MEGA X and manually trimmed before employing homology searches using BLASTn program against Gen Bank data base in NCBI [<a href=\"#r-22\">22</a>]. The evolutionary history was inferred by using the Maximum Likelihood method and kimura 2-parameter model [<a href=\"#r-23\">23</a>]. Initial trees (s) for the heuristic search were obtained automatically by applying Neighbour-Join and Bio NJ logarithms to a matrix of pairwise distances estimated using the Maximum Composite Likelihood (MCL) approach, and then selecting the topology with superior long likelihood value. The analysis involved 19 nucleotide sequences and there was a total of 747 positions in the final dataset. Evolutionary analysis was conducted in MEGA X [<a href=\"#r-24\">24</a>]. Data sequences for<em> Fusarium oxysporum,</em> a closely related species was retrieved from the database. A phylogenetic tree was constructed using the data sequences and reference to maximum likelihood method thereby analyzing the phylogenetic affiliations of the mangrove fungal endophytes [<a href=\"#r-25\">25</a>]. Construction of the phylogenetic tree in MEGA X used defaulting parameters and bootstrap standards derived after 1000 replications.</p>\r\n\r\n<p> </p>\r\n\r\n<p><em>Antimicrobial data analysis</em><br />\r\nA descriptive analysis of antibacterial activities and of ethyl acetate and methanolic extracts of mangrove endophytic fungi was performed and reported as mean Standard Deviation (SD) using Microsoft Excel. The MICs of ethyl acetate crude extracts against <em>Staphylococcus aureus</em> and <em>Escherichia coli</em> were determined and reported as mean Standard Error (SE). One-way ANOVA was used to compare the zone of inhibition of the fungal extracts against the positive and negative controls using STATISTICA statistical software version 7. The homoscedasticity of variances to meet the requirements of the analysis of variance (ANOVA) test was confirmed using Levene’s test at p > 0.05 and significance difference was assigned at p < 0.05.</p>"
},
{
"section_number": 3,
"section_title": "RESULTS",
"body": "<p><strong>Molecular identification of mangrove fungal endophytes</strong><br />\r\nA total of 50 fungal isolates were originally obtained from twig cuttings of selected mangroves [<a href=\"#r-14\">14</a>]. Bands of sufficient thickness were observed from the agarose gel electrophoresis and PCR amplicons (<a href=\"#figure2\">Figure 2</a>) from PCR. The average range of DNA concentration was 244.8 μg ml<sup>-1</sup> which was within the normal range at between 244.8 and 549.9 μg ml<sup>-1</sup>. The absorbance of DNA at A260 /A280 ranged at between 1.94 and 2.08. BLAST analysis gave closely related species (<a href=\"#Table-1\">Table 1</a>). 7 out 9 mangrove fungal isolates identified had 100 % similarity with the GenBank species. The 9 fungal isolates identified in this study were grouped into 3 clusters belonging to the phylum Ascomycota and genus <em>Aspergillus</em>.<br />\r\nResults of phylogenetic analysis (<a href=\"#figure3\">Figure 3</a>) showed that first cluster comprised of 3 isolates namely, AmT2, SaT1 and SaT3 grouped under <em>Aspergillus </em>with a 100 % bootstrap support representing<em> Aspergillus oryzae</em> (KP278199.1), <em>A. flavus </em>(MG799220.1) and <em>A. nomius </em>(KX431672.1) (<a href=\"#figure3\">Figure 3</a>). The second cluster showed 2 isolates, CtT1 and AmM2 under the genus <em>Aspergillus</em> with 98.81 % and 98.93 % bootstrap support, respectively representing <em>A. aculeatus </em>(KC621081.1) and <em>A. brunneoviolaceus </em>(MW180951.1). The third cluster showed 4 isolates namely, SaT2, AmG1, SaT4 and RmM4 with a 100 % bootstrap support. The cluster was made of <em>A. niger </em>(MG659662.1),<em> A. awamori </em>(MG659600.1)<em>, A. tubingensis </em>(MW466769.1) and<em> A. welwitchiae </em>(MK841443.1)<strong><em>.</em></strong><br />\r\nPhylogenetic analysis resulted in the tree with the highest log likelihood (-2246.83) (<a href=\"#figure3\">Figure 3</a>). The percentage of trees in which the associated taxa are clustered together is shown next to the branches of the phylogenetic tree (<a href=\"#figure3\">Figure 3</a>). Using universal primers ITS1 and ITS4A, a total of 9 out of 18 mangrove fungal isolates were successfully amplified. BLAST searches revealed the identities of the mangrove fungal isolates as members of one genus, <em>Aspergillus</em>, and the phylum Ascomycota. <em>Aspergillus flavus, Aspergillus niger, Aspergillus nomius, Aspergillus tubingensis, Aspergillus oryzae,</em> <em>Aspergillus awamori, Aspergillus aculeatus, Aspergillus brunneoviolaceus, </em>and<em> Aspergillus welwitchiae</em>. The genus <em>Aspergillus</em> dominated in this study accounting for 50% of the isolates identified (<a href=\"#figure3\">Figure 3</a>).</p>\r\n\r\n<div id=\"Table-1\">\r\n<p><a href=\"https://jabet.bsmiab.org/table/178-1650449385-table1/\">Table-1</a><strong>Table 1.</strong> Identification of mangrove fungal isolates from coastal Kenya using NCBI BLAST. </p>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<div id=\"figure2\">\r\n<figure class=\"image\"><img alt=\"\" height=\"186\" src=\"/media/article_images/2023/03/25/178-1650449385-Figure2.jpg\" width=\"428\" />\r\n<figcaption><strong>Figure 2.</strong> PCR Amplicons (approximately 500 base pairs) of ITS region of endophytic fungi isolated from selected mangrove species of coastal Kenya.</figcaption>\r\n</figure>\r\n</div>\r\n\r\n<div id=\"figure3\">\r\n<figure class=\"image\"><img alt=\"\" height=\"485\" src=\"/media/article_images/2023/03/25/178-1650449385-Figure3.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 3.</strong> Phylogenetic tree of 9 unique fungal isolates rooted from <em>Fusarium oxysporum</em> from coastal Kenya.</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Preliminary screening</strong><br />\r\nPreliminary screening of mangrove fungal extracts by paper disk diffusion showed that 4 out of 18 fungal endophytes inhibited the growth of test pathogens, gram-positive <em>Staphylococcus aureus </em>and gram-negative <em>Escherichia coli</em>. The zones of inhibition recorded at 50 mg/ml were between 5.67 and 17.1 mm. Methanolic extracts of isolates SaT2 and RmM4 inhibited both <em>S. aureus</em> and <em>E. coli</em> growth (<a href=\"#Table-2\">Table 2</a>). Methanolic extracts of isolates SaT1 and AmT2 only inhibited <em>S. aureus</em> (9.2 ± 0.75) and (6.16 ± 4.36), respectively. Figure 4 showed that ethyl acetate crude extracts of SaT1 exhibited the highest activity against both <em>S. aureus </em>(17.1 ± 2.45) and <em>E. coli </em>(15.9 ± 2.45). For all the extracts, the inhibition diameters of the fungal extracts were lower (5.67-17.1mm) than those of the positive control (24.77 ± 1.26). Results of 1-way ANOVA showed no significant difference in inhibition zone diameters between <em>S. aureus</em> and<em> E. coli</em> (df = 1; f = 2.182; p = 0.151)<br />\r\nThe values of the means of inhibition diameters appended by different superscripts along a column were significantly different (p < 0.05). For instance, 9.2±0.75<sup>a</sup> significantly differed from 6.16±4.36<sup>b</sup>. The values of means whose inhibition diameters within respective columns are followed by a superscript of a similar lower-case letter such as a and ac or c and ac were not significantly different from each other (p > 0.05<em>). </em>Means with different superscripts along the same row are significantly different (p < 0.05) (<a href=\"#Table-2\">Table 2</a>).</p>\r\n\r\n<div id=\"figure4\">\r\n<figure class=\"image\"><img alt=\"\" height=\"245\" src=\"/media/article_images/2023/03/25/178-1650449385-Figure4.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 4. </strong>Mean inhibition diameters of ethyl acetate crude extracts against <em>Staphylococcus aureus</em>, and <em>Escherichia coli</em>.</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<div id=\"Table-2\">\r\n<p><a href=\"https://jabet.bsmiab.org/table/178-1650449385-table2/\">Table-2</a><strong>Table 2</strong><em><strong>. </strong>In vitro</em> antibacterial susceptibility tests of crude extracts of mangrove fungal endophytes (Mangrove fungal endophytes from coastal Kenya).</p>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Determination of minimum inhibition concentration</strong><br />\r\nThe minimum inhibition concentration (MIC) value for <em>E. coli </em>was higher (0.91 ± 0.05 mg/ml) than that of <em>S. aureus </em>(0.82 ± 0.05 mg/ml) (<a href=\"#Table-3\">Table 3</a>). This implies that <em>S. aureus </em>was more susceptible to inhibition by active secondary metabolites of mangrove endophytic fungi than <em>E. coli. </em>The MIC values of the positive control for both test pathogens were higher than those of the test pathogens, ranging from 1.05 ± 0.10 to 1.08 ± 0.10 mg/ml for both Gram-positive <em>S. aureus </em>(ATCC 25922) and Gram-negative <em>E. coli</em> (ATCC 25922), respectively. <em>The </em>MIC value of <em>E. coli </em>(0.91 ± 0.05) was higher than that of <em>S. aureus </em>(0.82 ± 0.052).</p>\r\n\r\n<div id=\"Table-3\">\r\n<p><a href=\"https://jabet.bsmiab.org/table/178-1650449385-table3/\">Table-3</a><strong>Table 3. </strong>Minimum inhibition concentration of ethyl acetate crude extracts of <em>Aspergillus flavus</em> (Mangrove fungal endophytes from coastal Kenya).</p>\r\n\r\n<p> </p>\r\n</div>"
},
{
"section_number": 4,
"section_title": "DISCUSSION",
"body": "<p>A previous study isolated and identified the mangrove fungal endophytes from the twigs of mangrove species of <em>Rhizophora mucronata</em>, <em>Avicenia marina</em>, <em>Ceriops tagal,</em> and <em>Sonneratia alba </em>collected from coastal Kenya [<a href=\"#r-14\">14</a>]. ITS sequences identified 9 isolates (50%) from the genus <em>Aspergillus</em> belonging to the phylum Ascomycota as observed in this present study. In a similar study [<a href=\"#r-21\">21</a>], a total of 78 fungal isolates were identified from ITS sequences, 75 of which represented the phylum Ascomycota and only 3 of the phylum Basidiomycota. The phylum Ascomycota is said to be the most common representative of endophytic fungal communities isolated using standard procedures. It has also been reported that only a small portion of microorganisms in nature are culturable using standard microbiological techniques. This is because factors unfavorable for the growth of microorganisms exist in most laboratories. Some of the factors include limited knowledge of appropriate nutritive requirements and the challenging nature of microbes, especially when they are obtained from environmental samples [<a href=\"#r-21\">21</a>]. These findings are in agreement with the findings of the present study, where 9 (50%) of the 18 fungal isolates were fully identified as belonging to the genus <em>Aspergillus </em>in the family Ascomycota. It is thought that laboratory conditions may have interfered with the growth of the fugal isolates, only favoring the growth of <em>Aspergillus </em>sp. [<a href=\"#r-21\">21</a>]. <em>Aspergillus flavus </em>isolated from the mangrove plant <em>Avicenia officinalis </em>from Matang Mangrove Forest Reserve (MMFR), Perak, Malaysia was found to possess antioxidant properties [<a href=\"#r-21\">21</a>]. All the fungal isolates identified except isolate CtT1 and AmM2 identified as <em>Aspergillus aculeatus </em>and<em> Aspergillus brunneoviolaceus</em> showed a similarity of 98.81% and 98.93%, respectively, above the percentage identity threshold set at about 97.5%.<br />\r\nMangrove fungal endophytes have been a source of biologically active secondary metabolites with unique physiological and biochemical properties [<a href=\"#r-9\">9</a>, <a href=\"#r-26\">26</a>]. In the current study, both ethyl acetate and methanolic crude extracts of fungal endophytes from stem twigs of selected mangrove species of coastal Kenya showed significant resistance against standard strains of <em>S</em>.<em> aureus and E. coli</em>. This suggests that crude extracts of mangrove fungal endophytes contain biologically active compounds inhibiting the growth of both gram-positive <em>S. aureus </em>(ATCC 25922) and gram-negative <em>E. coli</em> (ATCC 25922) [<a href=\"#r-21\">21</a>] has reported the antibacterial activities exhibited by mangrove fungal extracts from the leaves of <em>Rhizophora mucronata</em> and <em>Sonneratia</em> <em>caseolaris</em>. In a similar study of antibacterial activities, ethyl acetate leaf extracts of different parts of <em>Avicenia marina</em> showed antibacterial activity against <em>S. aureus</em> and <em>E. coli </em>[<a href=\"#r-1\">1</a>]. The activity of mangrove endophytic fungal extracts was attributed to the presence of secondary metabolites such as alkaloids, flavonoids, quinones, and terpenoids. These extracts showed significant inhibition against multidrug pathogenic bacteria, <em>S. aureus</em> [<a href=\"#r-21\">21</a>]. In each case, inhibition diameters for both ethyl acetate and methanolic extracts were higher for <em>S. aureus </em>than for <em>E. coli. </em>This implies that the antibacterial activities of the fungal isolates were more common against the gram-positive <em>S. aureus</em> than the gram-negative <em>E. coli </em>and that<em> S. aureus</em> was more susceptible to the endophytic fungal extracts than <em>E. coli. </em>This observation was also reported [<a href=\"#r-27\">27</a>]. These authors stated that the higher resistance of gram-negative bacteria <em>E.</em> <em>coli</em> was attributed to the presence of an outer double membrane surrounding the cells, which prevents certain drugs and bioactive/antimicrobial compounds from penetrating the cells, thereby making <em>E</em>.<em> coli</em> more resistant than <em>S. aureus</em>. Generally, ethyl acetate crude extracts exhibited higher inhibition diameters than methanolic extracts. For instance, ethyl acetate crude extract of isolate SaT1 identified as <em>Aspergillus flavus</em> showed the highest activity against <em>S. aureus </em>(ATCC 25922) (17.1 mm) and <em>E</em>.<em> coli</em> (ATCC 25922) (15.9 mm) as compared to methanolic extracts, recording a maximum inhibition of 10.0 mm against<em> S. aureus </em>and 8.96 mm against <em>E. coli</em> at 50 g/ml<em>.</em><br />\r\nThe zones of inhibition of ethyl acetate extracts of <em>Avicenia marina</em> leaves by an <em>Aspergillus</em> sp. ranged from 8.33 to 10.67 mm at concentrations ranging from 60 to 100 mg/ml, according to [<a href=\"#r-27\">27</a>]. This is because leaf extracts are reported to contain more fungal metabolites than the stem twigs [<a href=\"#r-28\">28</a>].<em> A study </em>[<a href=\"#r-10\">10</a>] observed that ethyl acetate is preferred in the extraction of fungal metabolites because it is able to extract both extracellular fungal metabolites (excreted by fungi into the medium) and intracellular bioactive compounds, an observation that is in agreement with the findings of the current study. According to a study [<a href=\"#r-18\">18</a>], ethyl acetate is commonly used for the extraction of mangrove fungal metabolites since it is a medium-polar solvent with the ability to dissolve both polar and non-polar active compounds. This may imply that the most active secondary metabolites are medium polar and extractable by ethyl acetate. Methanolic crude extracts of all the fungal isolates showed activity against <em>S. aureus. </em>This implies that methanol was effective in extracting bioactive compounds from all the fungal endophytes that were active against <em>S. aureus</em> compared to ethyl acetate, an indicator that the activity of fungal extracts in this study was mainly as a result of polar active compounds extractable by methanol. According to a study [<a href=\"#r-18\">18</a>], because methanol is polar, it can dissolve almost all organic compounds, including polar, semi-polar, and non-polar compounds. <em>Aspergillus sp. </em>has been shown to be effective in inhibiting microbial growth [<a href=\"#r-21\">21</a>].<em> </em>In the present study, the most active fungal endophyte was found to be SaT1 <em>Aspergillus</em> <em>flavus</em> isolated from <em>A. marina</em> of Mida Creek, north coast Kenya [<a href=\"#r-14\">14</a>].<br />\r\nIn conclusion, the present study has revealed that selected mangrove species of coastal Kenya are rich in endophytic fungi which produce antibacterial substances. A total of 9 isolates were identified as belonging to one phylum, Ascomycota, and represented only one genus, <em>Aspergillus</em> sp. Two fungal species, <em>A. aculeatus </em>(MZ314732) and <em>A. brunneoviolaceus </em>(MZ314735), were identified with up to 98.81% and 98.93%, respectively, an observation that could suggest a need for further investigation. The findings of this study also confirm that endophytic fungi from harsh environments such as mangrove ecosystems are an attractive source of biologically active compounds inhibiting the growth of both gram-positive <em>S. aureus </em>(ATCC 25922) and gram-negative <em>E. coli</em> (ATCC 25922). Thu<em>s, </em>ethyl acetate crude extract of <em>Aspergillus</em> <em>flavus</em> showed greater activity against both test pathogens as compared to methanolic crude extracts.<br />\r\nEthyl acetate and methanol were used as extracting solvents in this study. The study recommends the use of other extracting solvents and subsequent tests against <em>S. aureus </em>(ATCC 25922) and <em>E. coli.</em> (ATCC 25922). This study also recommends further investigation of the rest of the mangrove fungal extracts for activity against a wider range of pathogenic microbes, which are largely underexplored in this region. Two of the fungal endophytes, <em>A. aculeatus </em>(MZ314732) and <em>A. brunneoviolaceus, </em>(MZ314735) were identified up to 98.81 and 98.93 (%), respectively, were not fully identified indicative of the need for further research on these two isolates. The isolation of the specific chemical agents responsible for activity against bacteria is hereby recommended for drug development and manufacture.</p>"
},
{
"section_number": 5,
"section_title": "ACKNOWLEDGEMENTS",
"body": "<p>The authors of this study appreciate the Technical University of Mombasa (TUM) and Kenya Marine and Fisheries Institute (KMFRI) for providing technical support and laboratory space over the study period. We also acknowledge Alexander Fulanda and Mackmillan for support in the isolation, purification, microscopy, antimicrobial assays, and morphological and molecular characterization of mangrove fungal endophytes at KMFRI laboratories. This study was partly funded by a marine research grant from the Western Indian Ocean Marine Science Association (WIOMSA) and the National Research Fund (NRF). It was also approved by the National Commission for Science, Technology, and Innovation (NACOSTI), (approval number: TUM ERC PHD/002/2020). This work forms part of the requirement for the Doctor of Philosophy Degree at TUM by the first author.</p>"
},
{
"section_number": 6,
"section_title": "AUTHORS CONTRIBUTION",
"body": "<p>HMK: Funding acquisition, Conceptualization, methodology, fieldwork, laboratory work, analysis and writing of original draft. CMK: Methodology, supervision, data analysis and editing; CNM: Funding acquisition, supervision, data analysis and editing; JOO: Fund acquisition, conceptualization, methodology, supervision and editing; PMG: Conceptualization, methodology, supervision and editing, and SMM: laboratory work and data analysis. All authors approved the final manuscript for publication.</p>"
},
{
"section_number": 7,
"section_title": "CONFLICTS OF INTEREST",
"body": "<p>There is no conflict of interest among the authors.</p>"
}
],
"figures": [
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/03/25/178-1650449385-Figure1.jpg",
"caption": "Figure 1. A map of the Kenya coast showing mangrove forests of Gazi Bay, Tudor and Mida creeks from where mangrove twig samples were collected [14].",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/03/25/178-1650449385-Figure2.jpg",
"caption": "Figure 2. PCR Amplicons (approximately 500 base pairs) of ITS region of endophytic fungi isolated from selected mangrove species of coastal Kenya.",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/03/25/178-1650449385-Figure3.jpg",
"caption": "Figure 3. Phylogenetic tree of 9 unique fungal isolates rooted from Fusarium oxysporum from coastal Kenya.",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/03/25/178-1650449385-Figure4.jpg",
"caption": "Figure 4. Mean inhibition diameters of ethyl acetate crude extracts against Staphylococcus aureus, and Escherichia coli.",
"featured": false
}
],
"authors": [
{
"id": 527,
"affiliation": [
{
"affiliation": "School of Applied and Health Sciences, Technical University of Mombasa, P.O. Box 90420 – 80100, Mombasa, Kenya"
}
],
"first_name": "Helen M.",
"family_name": "Kiti",
"email": "kitihelenmwaka@tum.ac.ke",
"author_order": 1,
"ORCID": "http://orcid.org/0000-0002-5287-3482",
"corresponding": true,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "Helen M. Kiti, PhD; School of Applied and Health Sciences, Technical University of Mombasa, Mombasa, Kenya e-mail: kitihelenmwaka@tum.ac.ke",
"article": 129
},
{
"id": 528,
"affiliation": [
{
"affiliation": "School of Applied and Health Sciences, Technical University of Mombasa, P.O. Box 90420 – 80100, Mombasa, Kenya"
}
],
"first_name": "Cromwell M.",
"family_name": "Kibiti",
"email": null,
"author_order": 2,
"ORCID": "http://orcid.org/0000-0002-3611-4427",
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 129
},
{
"id": 529,
"affiliation": [
{
"affiliation": "School of Applied and Health Sciences, Technical University of Mombasa, P.O. Box 90420 – 80100, Mombasa, Kenya"
}
],
"first_name": "Cosmas N.",
"family_name": "Munga",
"email": null,
"author_order": 3,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 129
},
{
"id": 530,
"affiliation": [
{
"affiliation": "School of Applied and Health Sciences, Technical University of Mombasa, P.O. Box 90420 – 80100, Mombasa, Kenya"
}
],
"first_name": "Josiah O.",
"family_name": "Odalo",
"email": null,
"author_order": 4,
"ORCID": "http://orcid.org/0000-0002-4032-4312",
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 129
},
{
"id": 531,
"affiliation": [
{
"affiliation": "Department of Chemistry, Pwani University, P.O. Box 195-80108, Kilifi, Kenya"
}
],
"first_name": "Paul M.",
"family_name": "Guyo",
"email": null,
"author_order": 5,
"ORCID": "http://orcid.org/0000-0002-3444-7973",
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 129
},
{
"id": 532,
"affiliation": [
{
"affiliation": "Kenya Marine and Fisheries Research Institute, P.O Box 81651-80100, Mombasa, Kenya"
}
],
"first_name": "Samuel M.",
"family_name": "Mwamburi",
"email": null,
"author_order": 6,
"ORCID": "http://orcid.org/0000-0003-4253-3967",
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 129
}
],
"views": 1181,
"downloads": 154,
"references": [
{
"id": 4279,
"serial_number": 1,
"pmc": null,
"reference": "Okla MK, Alatar AA, Al-Amri SS, Soufan WH, Ahmad A, Abdel-Maksoud MA. Antibacterial and Antifungal Activity of the Extracts of Different Parts of Avicennia marina (Forssk.) Vierh. Plants (Basel). 2021; 10(2).",
"DOI": null,
"article": 129
},
{
"id": 4280,
"serial_number": 2,
"pmc": null,
"reference": "Organization WH. Antimicrobial resistance. https://www.who.int/news-room/fact-sheets/detail/antimicrobial-resistance, 2021.",
"DOI": null,
"article": 129
},
{
"id": 4281,
"serial_number": 3,
"pmc": null,
"reference": "aranraj PSD, Mangrove Medicinal Plants: A Review. American-Eurasian Journal of Toxicological Sciences, 2015; 7(3): 146-156.",
"DOI": null,
"article": 129
},
{
"id": 4282,
"serial_number": 4,
"pmc": null,
"reference": "Dadar M, Tiwari R, Karthik K, Chakraborty S, Shahali Y, Dhama K. Candida albicans – Biology, molecular characterization, pathogenicity, and advances in diagnosis and control – An update. Microbial pathogenesis, 2018; 117: 128-138.",
"DOI": null,
"article": 129
},
{
"id": 4283,
"serial_number": 5,
"pmc": null,
"reference": "eoh WP, Chen X, Laczkovich I, Alonzo F. Staphylococcus aureus adapts to the host nutritional landscape to overcome tissue-specific branched-chain fatty acid requirement. Proceedings of the National Academy of Sciences of the United States of America, 2021; 118(13).",
"DOI": null,
"article": 129
},
{
"id": 4284,
"serial_number": 6,
"pmc": null,
"reference": "Chigodi MO. Isolation and screening of Antipneumonic compounds from Acacia Stuhlmannii Taub. (Master of Science in Chemistry pdf), Pwani University, 2017.",
"DOI": null,
"article": 129
},
{
"id": 4285,
"serial_number": 7,
"pmc": null,
"reference": "Huttanus HM, Sheng J, Feng X. Metabolic Engineering for Production of Small Molecule Drugs: Challenges and Solutions. Fermentation, 2016; 2(1): 4.",
"DOI": null,
"article": 129
},
{
"id": 4286,
"serial_number": 8,
"pmc": null,
"reference": "Bijaya KN. Studies on endophytic fungal diversity from different leaf samples of Pongamia pinnata. International Journal of MediPharm Research, 2015; 01(02): 134-138.",
"DOI": null,
"article": 129
},
{
"id": 4287,
"serial_number": 9,
"pmc": null,
"reference": "Cadamuro RD, da Silveira Bastos IMA, Silva IT, da Cruz ACC, Robl D, Sandjo LP, Alves SJr, Lorenzo J M, Rodriguez-Lazaro D, Treichel H, Steindel M, Fongaro G. Bioactive Compounds from Mangrove Endophytic Fungus and Their Uses for Microorganism Control. Journal of Fungi (Basel), 2021; 7(6).",
"DOI": null,
"article": 129
},
{
"id": 4288,
"serial_number": 10,
"pmc": null,
"reference": "Ling MO, Teen PL, Mujahidin A, Proksch P, Muller M. Initial Screening of Mangrove Endophytic Fungi for Antimicrobial Compounds and Heavy Metal Biosorption Potential. Sains Malaysiana, 2016; 45(7): 1063–1071.",
"DOI": null,
"article": 129
},
{
"id": 4289,
"serial_number": 11,
"pmc": null,
"reference": "Ling M, Onn L, Teen P, Aazani M, Peter P, Moritz, M. Initial screening of mangrove endophytic fungi for antimicrobial compounds and heavy metal biosorption potential. University of Malaya, 2013.",
"DOI": null,
"article": 129
},
{
"id": 4290,
"serial_number": 12,
"pmc": null,
"reference": "Programme, U. N. E. (2014). The importance of mangroves to people: a call to action. https://wedocs.unep.org/20.500.11822/9300.",
"DOI": null,
"article": 129
},
{
"id": 4291,
"serial_number": 13,
"pmc": null,
"reference": "Dahdouh-Guebas F, Mathenge C, Kairo JG, Koedam N. Utilization of mangrove wood products around Mida creek (Kenya) amongst subsistence and commercial users. Economic Botany, 2000; 54(4): 513-527.",
"DOI": null,
"article": 129
},
{
"id": 4292,
"serial_number": 14,
"pmc": null,
"reference": "Kiti HM, Munga CM, Odalo JO, Guyo PM, Kibiti CM. Diversity of mangrove fungal endophytes from selected mangrove species of coastal Kenya. Western Indian Ocean Journal of Marine Science, 2021; 20(1): 125-136.",
"DOI": null,
"article": 129
},
{
"id": 4293,
"serial_number": 15,
"pmc": null,
"reference": "Kairo JG, Dahdouh-Guebas F, Bosire J, Koedam N. Restoration and management of mangrove systems — a lesson for and from the East African region. South African Journal of Botany, 2001; 67(3): 383-389.",
"DOI": null,
"article": 129
},
{
"id": 4294,
"serial_number": 16,
"pmc": null,
"reference": "Jenoh EM, de Villiers EP, de Villiers SM, Okoth S, Jefwa J, Kioko E, Kaimenyi D, Hendrickx M, Dahdouh-Guebas F, Koedam N. Infestation mechanisms of two woodborer species in the mangrove Sonneratia alba J. Smith in Kenya and co-occurring endophytic fungi. PLOS ONE, 2019; 14(10): e0221285.",
"DOI": null,
"article": 129
},
{
"id": 4295,
"serial_number": 17,
"pmc": null,
"reference": "Bosire JO, Kaino JJ, Olagoke AO, Mwihaki LM, Ogendi GM, Kairo JG, Berger U, Macharia D. Mangroves in peril: unprecedented degradation rates of peri-urban mangroves in Kenya. Biogeosciences, 2014; 11(10): 2623-2634.",
"DOI": null,
"article": 129
},
{
"id": 4296,
"serial_number": 18,
"pmc": null,
"reference": "Nurunnabi T, Al-Majmaie S, Nakouti I, Nahar L, Rahman S, Sohrab M, Billah M, Ismail F, Sharples G, Sarker S. Antimicrobial activity of kojic acid from endophytic fungus Colletotrichum gloeosporioides isolated from Sonneratia apetala, a mangrove plant of the Sundarbans. Asian Pacific Journal of Tropical Medicine, 2018; 11(5): 350-354.",
"DOI": null,
"article": 129
},
{
"id": 4297,
"serial_number": 19,
"pmc": null,
"reference": "Kibiti CM, Afolayan AJ. Preliminary Phytochemical Screening and Biological Activities of Bulbine abyssinica Used in the Folk Medicine in the Eastern Cape Province, South Africa. Evidence-based complementary and alternative medicine: eCAM, 2015; 617607-617607.",
"DOI": null,
"article": 129
},
{
"id": 4298,
"serial_number": 20,
"pmc": null,
"reference": "Moron LS, Lim YW, Cruz TE. Antimicrobial activities of crude culture extracts from mangrove fungal endophytes collected in Luzon Island, Philippines, 2018.",
"DOI": null,
"article": 129
},
{
"id": 4299,
"serial_number": 21,
"pmc": null,
"reference": "Hamzah TNT, Lee SY, Hidayat A, Terhem R, Faridah-Hanum I, Mohamed R. Diversity and Characterization of Endophytic Fungi Isolated from the Tropical Mangrove Species, Rhizophora mucronata, and Identification of Potential Antagonists Against the Soil-Borne Fungus, Fusarium solani. Front Microbial, 2018; 9: 1707.",
"DOI": null,
"article": 129
},
{
"id": 4300,
"serial_number": 22,
"pmc": null,
"reference": "Thorati MMJK, Kumar S. Isolation, Identification of Endophytic Fungi from Mangrove Roots along the Coast of South Andaman Sea, Andaman and Nicobar Islands, India. Journal of Marine Biology & Oceanography, 2016; 5(2): 1-5.",
"DOI": null,
"article": 129
},
{
"id": 4301,
"serial_number": 23,
"pmc": null,
"reference": "Kimura M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. Journal of Molecular Evolution, 1980; 16(2): 111-120.",
"DOI": null,
"article": 129
},
{
"id": 4302,
"serial_number": 24,
"pmc": null,
"reference": "Kumar S, Kaushik N, Proksch P. Identification of antifungal principle in the solvent extract of an endophytic fungus Chaetomium globosum from Withania somnifera. SpringerPlus, 2013; 2(1): 37.",
"DOI": null,
"article": 129
},
{
"id": 4303,
"serial_number": 25,
"pmc": null,
"reference": "Mao Z, Zhang W, Wu C, Feng H, Peng Y, Shahid H, Cui Z, Ding P, Shan T. Diversity and antibacterial activity of fungal endophytes from Eucalyptus exserta. BMC Microbial, 2021; 21(1): 155.",
"DOI": null,
"article": 129
},
{
"id": 4304,
"serial_number": 26,
"pmc": null,
"reference": "Ababutain IM, Aldosary SK, Aljuraifani AA, Alghamdi AI, Alabdalall AH, Al-Khaldi EM, Aldakeel SA, Almandil NB, AbdulAzeez S, Borgio JF. Identification and Antibacterial Characterization of Endophytic Fungi from Artemisia sieberi. International journal of microbiology, 2021; 6651020-6651020.",
"DOI": null,
"article": 129
},
{
"id": 4305,
"serial_number": 27,
"pmc": null,
"reference": "Ariole CN, Akinduyite AE. Antibacterial potential of indigenous red mangrove (Rhizophora racemosa) fungal endophytes and bioactive compounds identification. International Network for Natural Sciences, 2016; 4: 14-24.",
"DOI": null,
"article": 129
},
{
"id": 4306,
"serial_number": 28,
"pmc": null,
"reference": "Ariole CN, Akinduyite, AE. Bioactive compounds and antimicrobial activity of endophytic fungi isolated from Black Mangrove (Avicenia Africana) leaves. Nigerian. Journal of Biotechnology, 2018; 35(2): 35 – 42.",
"DOI": null,
"article": 129
}
]
},
{
"id": 122,
"slug": "178-1652412212-spatial-distribution-of-phytoplankton-in-lasolo-bay-of-southeast-sulawesi-province-indonesia",
"featured": false,
"slider": false,
"issue": "Vol5 Issue3",
"type": "original_article",
"manuscript_id": "178-1652412212",
"recieved": "2022-05-13",
"revised": null,
"accepted": "2022-06-28",
"published": "2022-07-08",
"pdf_file": "https://jabet.bsmiab.org/media/pdf_file/2023/35/178-1652412212.pdf",
"title": "Spatial distribution of phytoplankton in Lasolo Bay of Southeast Sulawesi province, Indonesia",
"abstract": "<p>This study aimed to analyze the distribution of phytoplankton abundance and its relationship with water quality in Lasolo Bay waters. The study was conducted during the West season 2020. The sample consisted of 9 stations located near the mouth of the Lasolo River (station A), coastal waters (station B) and the center part of Lasolo Bay (station C). Phytoplankton sampling was using plankton nets with 25 µm mesh size. Identification of phytoplankton used microscope and the abundance calculation by using Sedgwick Rafter Counting Cell. The results showed that there are 23 phytoplankton genera from 5 classes found, namely 12 genera of <em>Bacillariophycea</em> class, 5 genera of <em>Dinophyceae</em> class, 3 genera of <em>Chlorophyceae</em> class, 2 genera of <em>Cyanophyceae</em> class and 1 genera of <em>Coscinodiscophyceae</em> class. By abundance, phytoplankton compositions of the class <em>Bacillariophyceae</em> predominate with a percentage of 57%. The type of phytoplankton that has the highest abundance is <em>Nitzschia</em> sp. of 2,407 cells/l and the lowest abundance of <em>Amphidium </em>sp. of 1,368 cells/l. Phytoplankton abundance at station A is 41,093 cells/l, station B is 14,234 cells/l and station C is 14,735 cells/l. The distribution of phytoplankton abundance is also influenced by physical and chemical factors such as turbidity, TSS, Nitrate, and Phosphate.</p>",
"journal_reference": "J Adv Biotechnol Exp Ther. 2022; 5(3): 615-622.",
"academic_editor": "Akhi Moni, PhD; ABEx Bio-Research Center, Dhaka-1230, Bangladesh",
"cite_info": "Irawati M, Sara L, et al. Spatial distribution of phytoplankton in Lasolo Bay of Southeast Sulawesi province, Indonesia. J Adv Biotechnol Exp Ther. 2022; 5(3): 615-622.",
"keywords": [
"Aquatic",
"Abundance",
"Phytoplankton"
],
"DOI": "doi.org/10.5455/jabet.2022.d140",
"sections": [
{
"section_number": 1,
"section_title": "INTRODUCTION",
"body": "<p>Lasolo Bay is one of the waters located in the North Konawe Regency. This area has aquatic resources and life support functions that have been designated as a nature conservation area and serve as a marine nature park [<a href=\"#r-1\">1</a>]. Nickel mining activities are carried out on land and Losolo beach [<a href=\"#r-2\">2</a>]. Mining activities such as dredging, laterite soil extraction, and transportation of minerals are placed along the coast. The entry of mining waste materials through the surface flow in the Lasolo river basin causes pollution in the Lasolo Bay area [<a href=\"#r-3\">3</a>]. Turbid water conditions cause some coral reefs to have been covered with mud. The occurrence of turbidity in the waters causes reduced light penetration in seawater which can affect the growth and development of aquatic organisms. One of the marine organisms that is important for the productivity of a body of water is phytoplankton. Phytoplankton is a major producer that is able to produce energy through the process of photosynthesis [<a href=\"#r-4\">4</a>]. Phytoplankton abundance is one indicator of the quality and productivity of aquatic ecosystems [<a href=\"#r-5\">5</a>]. Distribution of phytoplankton abundance depends on the condition of the aquatic environment.<br />\r\nChanges in water quality can be viewed from the composition and abundance of phytoplankton. Hydrodynamic conditions of an aquatic will affect the distribution pattern of phytoplankton horizontally and vertically. Research on phytoplankton has never been conducted in these waters. Information about the composition and abundance of phytoplankton is essential to determine the productivity of waters</p>"
},
{
"section_number": 2,
"section_title": "MATERIALS AND METHODS",
"body": "<p><strong>Sampling locations and procedures</strong><br />\r\nThis study was conducted in December 2020 to coincide with the Western monsoon in the waters of Lasolo Bay located at coordinates 122<sup>0</sup>12’00<sup>’’ </sup>– 122<sup>0</sup>27’00<sup>’’</sup>BT and 3<sup>0</sup>30’ 00<sup>’’</sup> – 3<sup>0</sup>45’00<sup>’’</sup>LS. Sampling was carried out in 3 zones, zone A is located in front of the mouth of the Lasolo River, zone B is located on the coast, and zone C is located in the middle of Lasolo Bay. Each zone consists of 3 stations, so a total of 9 stations (<a href=\"#figure1\">Figure 1</a>). Vertically, each station is divided into 3 station points based on the depth of sunlight intensity in the water column, namely at a depth of light intensity of 100%, 50%, and 25% of the light intensity of the water surface. Sampling is done horizontally and vertically. Samples collected include the waters’ physical, chemical, and biological parameters (temperature, pH, salinity, brightness, and turbidity). Total Suspended Solid (TSS), Nitrate, Orthophosphate, and phytoplankton abundance were also included. Furthermore, sample collection and measurement were repeated at each station (A, B, and C).</p>\r\n\r\n<div id=\"figure1\">\r\n<figure class=\"image\"><img alt=\"\" height=\"220\" src=\"/media/article_images/2023/29/26/178-1652412212-Figure1.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 1. </strong>Lasolo Bay location map.</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Phytoplankton sampling</strong><br />\r\nWater sampling horizontally using Nansen bottles taken on the surface of the waters and at a depth of sunlight penetration intensity of 50% and 25% from the surface. Determination of the depth of sunlight intensity based on the Beer-Lambert law [<a href=\"#r-6\">6</a>]<br />\r\n<em>Formula</em>………………………………. (1)<br />\r\nWhere, <em>Iz</em> = Light intensity at depth<em> z</em>, <em>Io</em> = Light intensity on the surface of the waters, e = Natural logarithm (2.70), <em>k</em> = Dimming coefficient, and z = Depth<br />\r\nVertical water intake is carried out using a pump connected with a hose to the surface of the water. Furthermore, the sample was filtered using a plankton net with a diameter of 31 cm with a mesh size of 25μm. The sample was stored in a 100 ml vial and preserved using 4% acetic acid lugol. When taking water samples horizontally and vertically, measurements of water chemical physics parameters are also carried out, namely brightness measurements carried out using a secchi disk.<br />\r\nTemperature and salinity measurements use a Hg thermometer (-10 – 110 <sup>0</sup>C) and a hand fefractometer alcohol ATC. Sea water pH measurement uses a pH meter (Pen type PH-009 I A). Measurement of Nitrates and Phosphates (Orthophosphates) using the standard method of Spectrophotometers. Turbidity is measured in the laboratory based on the Nephelometric method to determine the TSS first, the sample was filtered using filter paper (Whatman) pore size of 0.45 mm, then the sample was analyzed using gravimetric methods.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Phytoplankton analysis</strong><br />\r\nThe composition and abundance of phytoplankton were analyzed by the Sedgewick Rafter Counting Cell (SRCC) method using equation [<a href=\"#r-7\">7</a>]; <em> Formula</em>……………………………. (2)<br />\r\nWhere <em>N </em>is the abundance of phytoplankton (cells/l), <em>n</em> is the number of cells that are chopped, <em>V<sub>d</sub></em> is the volume of the water sample to be filtered (ml), <em>V<sub>t</sub> </em>is the volume of the filtered water sample (ml), and <em>V</em><em><sub>cg</sub></em> is the volume of the Sedwick Rafter Cell (ml).<br />\r\nThe sample was observed under a microscope (stereo zoom SZX7) with a magnification of 10×40 based on the sweep method, that is, counting all types of plankton present in the volume of the water sample. Phytoplankton is identified using plankton identification manuals. Phytoplankton was identified using the plankton identification manual [<a href=\"#r-8\">8</a>]. The results of the calculations are further analyzed to find out the composition and abundance of phytoplankton.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Statistical analysis</strong><br />\r\nThe data obtained were analyzed using Excel data sheets to determine the correlation between phytoplankton abundance and aquatic chemical physics parameters. The data were analyzed using multiple linear regression analysis and F-test. This test was carried out to see the relationship between chemical physics parameters to phytoplankton abundance.</p>"
},
{
"section_number": 3,
"section_title": "RESULTS",
"body": "<p><strong>Phytoplankton type composition.</strong><br />\r\nIn the Lasolo Bay waters, we found 23 genera of 5 phytoplankton classes consisting of 12 genera the <em>Bacillariophyceae</em> class, 5 genera the <em>Dinophyceae</em> class, 3 genera the <em>Chlorophyceae</em> class, 2 genera the <em>Cyanophyceae</em> class, and 1 genera the <em>Coscinodiscophyceae</em> class (<a href=\"#Table-1\">Table 1</a>). The abundance of phytoplankton at station A is greater than that of stations B and C. Types of phytoplankton that have the highest-class composition are <em>Bacillaryophyceae</em> with a percentage of 52%, while the classes <em>Dinophyceae, Chlorophyceae Cyanophyceae</em> and <em>Coscinodiscophyceae</em> are 22%, 13% 9% and 4%, respectively (<a href=\"#figure2\">Figure 2a</a>). The highest abundance was found in the class <em>Bacillaryophyceae</em> of 25,166 cells/l, with a percentage of 57%. Class <em>Coscinodiscophycea</em>e has the lowest abundance of 2,072 cells/l with a portion of 5% (<a href=\"#figure2\">Figure 2b</a>).</p>\r\n\r\n<div id=\"Table-1\">\r\n<p><a href=\"https://jabet.bsmiab.org/table/178-1652412212-table1/\">Table-1</a><strong>Table 1.</strong> Abundance of phytoplankton types.</p>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<div id=\"figure2\">\r\n<figure class=\"image\"><img alt=\"\" height=\"179\" src=\"/media/article_images/2023/30/26/178-1652412212-Figure2.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 2. </strong>(a) Phytoplankton class composition; (b) Abundance of phytoplankton.</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Abundance of phytoplankton</strong><br />\r\nThe distribution of phytoplankton abundance varied horizontally and vertically following the depth of sunlight penetration at each station. The highest abundance of phytoplankton between stations in the waters of Lasolo Bay was found at station A at 41,093 cells/l. The phytoplankton abundance at stations B and C was 14,234 cells/l and 14,735 cells/l, respectively. A graph of the abundance of types between stations can be seen in <a href=\"#figure3\">Figure 3</a>. The type of phytoplankton with the highest abundance is found at station A, namely <em>Amphora</em> sp. by 1,117 cells / l, at station B is <em>Microcystus</em> sp. by 1,017 cells/l and station C is <em>Rhizosolenia</em> sp. by 981 cells/l. In contrast, the lowest type of abundance is <em>Peridinium sp</em>. of 313 cells / l (<a href=\"#Table-1\">Table 1</a>).</p>\r\n\r\n<div id=\"figure3\">\r\n<figure class=\"image\"><img alt=\"\" height=\"290\" src=\"/media/article_images/2023/30/26/178-1652412212-Figure3.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 3. </strong>Graph of phytoplankton abundance at stations A, B, and C.</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Water physicochemical conditions</strong><br />\r\nThe physical and chemical characteristics of Lasolo Bay water are shown in <a href=\"#Table-2\">Table 2</a>.</p>\r\n\r\n<div id=\"Table-2\">\r\n<p><a href=\"https://jabet.bsmiab.org/table/178-1652412212-table2/\">Table-2</a><strong>Table 2.</strong> Water quality measurement.</p>\r\n\r\n<p> </p>\r\n</div>"
},
{
"section_number": 4,
"section_title": "DISCUSSION",
"body": "<p>The composition of phytoplankton by the number of species in the waters of Lasolo Bay is dominated by the class <em>Bacillariophyceae</em> (Diatoms). The class <em>Bacillariophyceae</em> dominates with a percentage of more than 52% because this phytoplankton is able to adapt to the environment and has a high reproductive power [8]. The distribution of phytoplankton with the highest abundance is found at station A with a range of 4,067 – 4,960 cells/l in front of the mouth of the Lasolo river. The average abundance at station A is present on the surface and decreases with increasing depth. The intensity of reduced sunlight (50% and 25%) may cause a lower abundance of phytoplankton [<a href=\"#r-9\">9</a>].<br />\r\nThe spatial distribution of the abundance of phytoplankton types varies at each station. The phytoplankton found are scattered almost over the entire surface of the waters, and the depth is suitable for sunlight penetration. Changes in abundance tend to decrease towards the coast towards the central part of the bay [<a href=\"#r-10\">10</a>]. <em>Bacillariophyceae</em> and <em>Cyanophyceae</em> have a higher abundance than other classes found. Some factors that can support the abundance of phytoplankton include the presence of nutrients such as Nitrates and Phosphates [<a href=\"#r-11\">11</a>]. The total abundance of phytoplankton on the surface (0 m) is 14,058 cells/l, at a depth of 0.5-2 m of 15,323 cells/l and a depth of 2 – 5 m of 14,887 cells/l.<br />\r\nVertically, the highest abundance was found at station A2.2 at a light intensity depth of 50% of the surface intensity or at a depth of 2.8 meters by 2021 cells/l. In comparison, the highest abundance of types is found at stations A1.1 and A2.3 with the most types of phytoplankton being <em>Amphora</em> sp., as many as 1,117 cells/l. The lowest abundance of the type is <em>Merismopedia</em> sp. of 312 cells/l. The <em>Bacillariophyceae</em> class dominates the phytoplankton composition at station B on the coast and has the highest number of types at various depths. The highest abundance at station B3.3 at a depth of 4.3 m is 1,751 cells/l and is dominated by the <em>Cyanophyceae</em> class, namely <em>Microcystus </em>sp., which is 1017 cells/l. At station C, the composition of phytoplankton is also dominated by the class <em>Bacillaryophyceae</em>, a type of <em>Rhizosolenia </em>sp. Further, classes are Dinophyceae (2,577 cells / l), Chlorophyceae (1996 cells/l), Cyanophyceae (873 cells/l), and Coscinodiscophyceae (707 cells/l).<br />\r\nThe difference in abundance is due to the influence of activities around the waters and conditions at each station. Each site has a different anthropogenic impact, and nutrient supply also affects the growth of phytoplankton in the waters [<a href=\"#r-12\">12</a>]. High nutrient levels are generally found on parts of the surface that are still affected by freshwater, such as at the mouth of rivers. The nutrient is derived from human activities on land [<a href=\"#r-13\">13</a>]. The higher abundance of phytoplankton at station A (Figure 3) is due to being influenced by nutrients coming from land that enter the waters through streams and surface flows.<br />\r\nEnvironmental factors also influence the distribution and growth rate of phytoplankton, among others temperature, salinity, turbidity, TSS, and nutrients. River estuaries cause the high abundance at station A with high nutrient levels, namely Phosphate 0.71 mg/l and Nitrate 0.52 mg/l. In addition, low turbidity and TSS also affect the distribution of phytoplankton abundance. Table 2 shows station A located at the mouth of the Lasolo river, has low turbidity and TSS concentration and conversely has a high abundance of phytoplankton. The concentration of TSS at station A was 140 mg/l – 420 mg/l with a higher phytoplankton abundance value of 41,093 cells /l. Stations B and C, have a turbidity value range of 13.5 mg/l – 28.1 mg/l, with the highest TSS value of 570 mg/l and the lowest of 290 mg/l. TSS concentration influences phytoplankton’s abundance; the lower the concentration, the greater the abundance of phytoplankton [<a href=\"#r-14\">14</a>].<br />\r\nSeawater temperature is also critical in metabolic processes and reproductive abilities affecting the composition and abundance of phytoplankton [<a href=\"#r-15\">15</a>]. The temperature range in the waters of Lasolo Bay ranges from 28.7 – 31.8 <sup>0</sup>C. The highest temperature is at station C3.1, and the lowest is at station A2.1. The temperature distribution shows the value is getting higher towards the middle of the bay because sampling is carried out in different timescales from morning to noon. However, the temperature range during the study was still within the temperature range corresponding to the growth and development of phytoplankton, namely 28-32 <sup>0</sup>C [<a href=\"#r-16\">16</a>].<br />\r\nThe salinity obtained at station A ranges from 2.6 – 30.6 o/oo because the station is located around the mouth of the Lasolo river, which is influenced by freshwater input from the mainland, especially at high tide and low tide. At station B the salinity ranges from 21.6 – 32.6 o/oo, and at station C it ranges from 16.5 – 31 o/oo. Generally, low salinity values are found near the mouth of the Lasolo river. The low salinity value is due to its location adjacent to the mouth of the Lasolo river which is strongly influenced by the tidal activity of the waters.<br />\r\nThe correlation between phytoplankton abundance and aquatic chemical physics parameters is based on correlation tests with a value of 0.743 and an F significance value of 0.016. These values indicate a strong correlation between phytoplankton abundance and water quality parameters. These physical and chemical factors cause the distribution of phytoplankton to be uneven so that they have varying abundances [<a href=\"#r-17\">17</a>]. Phytoplankton abundance values found at station A amounted to 41,093 cells/l, station B was 14,234 cells/l, and station C was 14,735 cells/l.</p>"
},
{
"section_number": 5,
"section_title": "CONCLUSION",
"body": "<p>In the Lasolo Bay waters found 23 genera of 5 phytoplankton classes consisting of 12 genera the Bacillariophyceae class, 5 genera the Dinophyceae class, 3 genera the Chlorophyceae class, 2 genera the Cyanophyceae class and 1 genus the Coscinodiscophyceae class. The highest abundance of phytoplankton was found at station A at 41,093 cells/l. and the lowest abundance was at station B of 14,234 cells/l. Based on the depth of the sunlight intensity entering the waters, the distribution of phytoplankton abundance with an intensity of 100%, 50% and 25% of the surface intensity was 14,058 cells/l, 15,323 cells/l, and 14,887 cells/l. The type of phytoplankton that has the highest abundance is Nitzschia sp. of 2,407 cells/l and the lowest abundance of Amphidium sp. of 1,368 cells/l. The distribution of the abundance of phytoplankton is also influenced by the physical and chemical factors of the waters, namely turbidity, TSS, Nitrate, and Phosphate. The correlation between the abundance of phytoplankton with the physical parameters of aquatic chemistry based on a correlation test with a value of 0.743 and the significance value of 0.016 shows a strong correlation.</p>"
},
{
"section_number": 6,
"section_title": "ACKNOWLEDGEMENTS",
"body": "<p>Thanks to Prof. Ir. H. La Sara, Ph.D., Dr. Muliddin and Prof. Dr. Asriyana, S.Pi.,M.Si for his guidance and direction in completing this research. I would also like to thank the Biology Laboratory of Halu Oleo University for its assistance in facilitating and analyzing water quality data.</p>"
},
{
"section_number": 7,
"section_title": "AUTHORS CONTRIBUTION",
"body": "<p>Irawati; Develop and design experiments, equipment, prepare materials, sample data, and analyze data. La Sara, Muliddin, Asriyana; designing experiments, guiding drafting manuscripts, correcting manuscripts, and providing technical input.</p>"
},
{
"section_number": 8,
"section_title": "CONFLICTS OF INTEREST",
"body": "<p>There is no conflict of interest among the authors.</p>"
}
],
"figures": [
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/29/26/178-1652412212-Figure1.jpg",
"caption": "Figure 1. Lasolo Bay location map.",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/30/26/178-1652412212-Figure2.jpg",
"caption": "Figure 2. (a) Phytoplankton class composition; (b) Abundance of phytoplankton.",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/30/26/178-1652412212-Figure3.jpg",
"caption": "Figure 3. Graph of phytoplankton abundance at stations A, B, and C.",
"featured": false
}
],
"authors": [
{
"id": 492,
"affiliation": [
{
"affiliation": "Doctoral Student of Agricultural Science, Postgraduate Program, Halu Oleo University, Kendari, Southeast Sulawesi, Indonesia"
}
],
"first_name": "Irawati",
"family_name": "M",
"email": "irawatijumaidin@gmail.com",
"author_order": 1,
"ORCID": null,
"corresponding": true,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "Irawati, Agricultural Science, Postgraduate Program, Halu Oleo University,\r\nKendari, Southeast Sulawesi, Indonesia, e-mail: irawatijumaidin@gmail.com",
"article": 122
},
{
"id": 493,
"affiliation": [
{
"affiliation": "Department of Aquatic Resources Management, Faculty of Fisheries and Marine Science, Halu Oleo University, Kendari, Southeast Sulawesi,Indonesia"
}
],
"first_name": "La",
"family_name": "Sara",
"email": null,
"author_order": 2,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 122
},
{
"id": 494,
"affiliation": [
{
"affiliation": "Department of Geological Engineering, Halu Oleo University, Kendari, Southeast Sulawesi, Indonesia"
}
],
"first_name": "Muliddin",
"family_name": "M",
"email": null,
"author_order": 3,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 122
},
{
"id": 495,
"affiliation": [
{
"affiliation": "Department of Aquatic Resources Management, Faculty of Fisheries and Marine Science, Halu Oleo University, Kendari, Southeast Sulawesi, Indonesia"
}
],
"first_name": "Asriyana",
"family_name": "M",
"email": null,
"author_order": 4,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 122
}
],
"views": 529,
"downloads": 113,
"references": [
{
"id": 4076,
"serial_number": 1,
"pmc": null,
"reference": "Kusmanto E, Surinati D. Stratifikasi massa air di Teluk Lasolo , Sulawesi Tenggara, Oseanologi dan Limnol. di Indonesia. 2016; 1(2): 17–29.",
"DOI": null,
"article": 122
},
{
"id": 4089,
"serial_number": 2,
"pmc": null,
"reference": "Ahmad F, “Dampak aktivitas perkotaan dan penambangan Nikel terhadap tingkat kontaminasi logam berat dalam air laut dan sedimen,” Indonesia. J. Mar. Sci. 2013; 18 (2): 71–78.",
"DOI": null,
"article": 122
},
{
"id": 4090,
"serial_number": 3,
"pmc": null,
"reference": "Setyawan WB, “Muatan sedimen tersuspensi dan kualitas lingkungan perairan Kendari, Sulawesi Tenggara,” 2016; 4: 495–501.",
"DOI": null,
"article": 122
},
{
"id": 4091,
"serial_number": 4,
"pmc": null,
"reference": "I. M. S. and Rissik D, Plankton. Australia: CSIRO , 2009.",
"DOI": null,
"article": 122
},
{
"id": 4092,
"serial_number": 5,
"pmc": null,
"reference": "Sahami FM, Baruadi ASR and Hamzah SN, “Proceeding the 1st International Seminar on sustainability in the marine fisheries sector 2017,” in Study of Phytoplankton Abundance for the Development of Pearl Oyster Culture Area in the North Gorontalo Waters, 2017; 1–7.",
"DOI": null,
"article": 122
},
{
"id": 4093,
"serial_number": 6,
"pmc": null,
"reference": "Asriyana and Yuliana, Produktivitas Perairan, Edisi revisi. Kendari: PT Bumi Aksara, 2019.",
"DOI": null,
"article": 122
},
{
"id": 4094,
"serial_number": 7,
"pmc": null,
"reference": "APHA, Standard Methods for the Examination of Water and Wastewater, 2nd ed. Washington DC: American Public Health Association/American Water Works Association/Water Environment Federation, 2005.",
"DOI": null,
"article": 122
},
{
"id": 4095,
"serial_number": 8,
"pmc": null,
"reference": "Odum EP, Ecology, Edisi 3. Yogyakarta: Gadjah Mada University Press Yogyakarta, 1993.",
"DOI": null,
"article": 122
},
{
"id": 4096,
"serial_number": 9,
"pmc": null,
"reference": "Muhtadi A, “Produktivitas Primer Perairan,” Researchgate.Net, 2017; 14(1): 1–19.",
"DOI": null,
"article": 122
},
{
"id": 4097,
"serial_number": 10,
"pmc": null,
"reference": "Ryabushko LI, Lishaev DN and Kovrigina NP, “Species diversity of epilithon diatoms and the quality of the waters of the Donuzlav gulf ecosystem (Crimea, the Black sea),” Diversity, 2019; 11( 7).",
"DOI": null,
"article": 122
},
{
"id": 4098,
"serial_number": 11,
"pmc": null,
"reference": "Aminah S, Nuraini RA, and Djunaedi A, “Komposisi dan kelimpahan fitoplankton di perairan Pandansari, Desa Kaliwlingi, Kabupaten Brebes, Jawa Tengah,” J. Mar. Res., 2020; 9(1): 81–86.",
"DOI": null,
"article": 122
},
{
"id": 4099,
"serial_number": 12,
"pmc": null,
"reference": "Wang X, Sun J and Yu H, “Distribution and environmental impact factors of phytoplankton in the Bay of Bengal during autumn,” Diversity, 2022; 14(5): 1–12.",
"DOI": null,
"article": 122
},
{
"id": 4100,
"serial_number": 13,
"pmc": null,
"reference": "Simanjuntak M, “Hubungan Faktor Lingkungan Kimia, Fisika terhadap Distribusi Plankton di Perairan Belitung Timur, Bangka Belitung,” J. Perikan., 2009; 11(1); 31–45.",
"DOI": null,
"article": 122
},
{
"id": 4101,
"serial_number": 14,
"pmc": null,
"reference": "Edje BO, Ishaque AB, and Chigbu P, “Spatial and temporal patterns of d13C and d15N of suspended particulate organic matter in Maryland Coastal Bays, USA,” Water (Switzerland), 2020; 12(9): 1–21.",
"DOI": null,
"article": 122
},
{
"id": 4102,
"serial_number": 15,
"pmc": null,
"reference": "Lo WT, Hsu PK, Fang TH, Hu JH, and Hsieh HY, “Phytoplankton Communities impacted by thermal effluents off two coastal nuclear power plants in subtropical areas of northern Taiwan,” Terr. Atmos. Ocean. Sci., 2016; 27(1): 107–120.",
"DOI": null,
"article": 122
},
{
"id": 4103,
"serial_number": 16,
"pmc": null,
"reference": "Hidup MNL, “Salinan Keputusan Menteri Negara Lingkungan Hidup Nomor: 51 Tahun 2004 Tentang Baku Mutu Air Laut,”. April, Jakarta:, 2004; (1–5).",
"DOI": null,
"article": 122
},
{
"id": 4104,
"serial_number": 17,
"pmc": null,
"reference": "Wulandari DY, Pratiwi NTM, and Adiwilaga EM , “Distribusi Spasial Fitoplankton di Perairan Pesisir Tangerang ( Spatial Distribution of Phytoplankton in the Coast of Tangerang ),” Ilmu Pertan. Indonesia., 2014; 19(3): 156–162.",
"DOI": null,
"article": 122
}
]
},
{
"id": 118,
"slug": "178-1653458316-protective-effect-of-l-carnitine-nanoparticles-vs-carnitine-on-lead-acetate-induced-toxicity-in-male-rats",
"featured": false,
"slider": false,
"issue": "Vol5 Issue3",
"type": "original_article",
"manuscript_id": "178-1653458316",
"recieved": "2022-05-25",
"revised": null,
"accepted": "2022-06-21",
"published": "2022-07-07",
"pdf_file": "https://jabet.bsmiab.org/media/pdf_file/2023/22/178-1653458316.pdf",
"title": "Protective effect of L-carnitine nanoparticles Vs carnitine on lead acetate-induced toxicity in male rats",
"abstract": "<p>Chitosan nanoparticles are important materials that are widely used in many biological, engineering and food industries and are also used as plant growth stimulants as well as use as vectors for drug delivery to target cells. Whereas L-carnitine (LC) is a water-soluble compound that contributes to the transport of long-chain fatty acids across the mitochondrial membranes and the oxidation of β-lipids. 60 male rats (Rattus Rattus) were divided into six equal groups. The first group (control group) received orally distilled water. The second group received 1ml lead acetate orally at a dose of 30 mg/kg of body weight daily for 30 days. Third group received 1ml lead acetate (30mg/kg B.W) + L-carnitine (100mg/ kg B.W. /daily). The fourth group received 1ml lead acetate (30mg/kg B.W.) + Nano L-carnitine (100mg/ kg B.W./ daily). The fifth group received 1ml of L-Carnitine orally at a dose of 100mg/ kg B.W /daily. The sixth group received 1ml of L-Carnitine-NPs orally at 100mg/ kg B.W /daily. Our findings demonstrated that exposure to lead acetate caused a significant increase in liver enzymes aspartate transaminase (AST), alanine transaminase (ALT), and alkaline phosphatase (ALP) and renal function (creatinine and urea) in the lead acetate group. Whereas lead treatment increased oxidative stress and reactive oxygen species (ROS). Histopathological study showed significant changes in the brain (cerebellum) that disrupted the normal arrangement of the three layers, with large distances between the Purkinje cell layer and the molecular or granular layer. According to the study, we can conclude that the Nano L-Carnitine had a greater role in protecting against the effect of lead at the hematological parameters and a clear role in the protection against histopathology change of lead poisoning. L-Carnitine and Nano L-Carnitine had an active role in protecting against lead acetate toxicity.</p>",
"journal_reference": "J Adv Biotechnol Exp Ther. 2022; 5(3): 590-604.",
"academic_editor": "Md Jamal Uddin, PhD; ABEx Bio-Research Center, Dhaka-1230, Bangladesh",
"cite_info": "Mahdi HT, Ghadhban RF, Protective effect of L-carnitine nanoparticles Vs carnitine on lead acetate-induced toxicity in male rats. J Adv Biotechnol Exp Ther. 2022; 5(3): 590-604.",
"keywords": [
"Liver",
"Nanoparticles",
"Chitosan",
"Kidney function",
"Brain",
"L-carnitine",
"Lead acetate"
],
"DOI": "10.5455/jabet.2022.d138",
"sections": [
{
"section_number": 1,
"section_title": "INTRODUCTION",
"body": "<p>In recent years, tremendous progress has been made in nanotechnology, particularly in the fields of material science and medicine. Medical applications of nanotechnology are often referred to as “nanomedicine.” This has provided a crucial impetus for creating several types of drug-loaded nanocarriers ranging in size from 1 to 1000 nm. In the biomedical field, a wide range of nanocarriers or nanoparticulate systems composed of various materials such as lipids, polymers, and inorganic materials have been proposed. That results in delivery systems depending on their physicochemical properties, which will be suitable for various applications [<a href=\"#r-1\">1,2</a>]. Chitosan is a linear polysaccharide formed from chitin, an abundant natural polymer found in crustaceans, insects, arthropods, and fungal cell walls. Most commercially available chitosan is generated from marine chitin collected from shrimp, lobster, and crab shells. Chitosan is the N-deacetylate form of chitin composed of D glucosamine and N-acetyl glucosamine monomers [<a href=\"#r-3\">3,4</a>].<br />\r\nChitosan advantages have shown that it may be used as a drug delivery system for the controlled release of antibiotics, anticancer medications, antihypertensive agents, proteins, peptide pharmaceuticals, and vaccines [<a href=\"#r-5\">5</a>]. Chitosan has also been used to increase the solubility of insoluble drugs in water, tissue engineering, ocular bandage lenses, gene delivery, and other uses are also possible [<a href=\"#r-6\">6,7</a>]. L-carnitine is a natural substance that prevents long-chain fatty acid accumulation by transporting them into the mitochondria and oxidizing them to produce adenosine triphosphate [<a href=\"#r-8\">8-10</a>]. L-carnitine suppresses both oxidative stress-induced mitochondrial damage and mitochondria-dependent apoptosis in various cell types [<a href=\"#r-11\">11</a>].<br />\r\nAccording to recent research, L-carnitine may play a significant role in oxidative/antioxidative equilibrium and has an anti-peroxidative impact on various tissues [<a href=\"#r-12\">12,13</a>]. Lead (Pb) is one of the most dangerous heavy metals found in the environment [<a href=\"#r-14\">14-16</a>]. Because lead has no good properties, its presence in the body in high quantities affects all animal organs, resulting in hazardous consequences [<a href=\"#r-17\">17</a>]. Lead exposure in the workplace and environment has grown many-fold, resulting in a wide range of uses in industries, cosmetics, folk cures, pharmaceuticals, and medicine. [<a href=\"#r-18\">18,19</a>]. Lead acetate is a bio-toxic contaminant that accumulates in all bodily tissues, including the liver, lungs, bones, reproductive systems, brain, and immune system. This hazardous lead’s physiological, biochemical, and behavioral consequences in animals have been recorded.<br />\r\nHigh in vivo stability, long-term payload capacity release and passage via small capillaries and cellular compartments are only a few of the various advantages of employing nanoparticle delivery systems. Nanoparticles may also increase the therapeutic index of the treatment, control its pharmacokinetics and biodistribution, and aid in the formation of long-term drug reservoirs [<a href=\"#r-7\">7</a>]. Additional characteristics include smaller nanoparticles (50-200 nm) with high loading capacity, delayed complex dissociation in vivo, and target optimization to the targeted location with limited absorption by neighboring tissues. Effective delivery methods need the development of formulations that include these features while being cost and complexity-effective [<a href=\"#r-17\">17</a>,<a href=\"#r-20\">20</a>].</p>"
},
{
"section_number": 2,
"section_title": "MATERIALS AND METHODS",
"body": "<p><strong>Chitosan-carnitine acid adduct (Cs-Ca) synthesis</strong><br />\r\nAccording to references, these steps were used to make chitosan-l-carnitine adduct [<a href=\"#r-21\">21</a>, <a href=\"#r-22\">22, 23</a>]. 1% of l-carnitine and chitosan were combined in equal amounts using Dean-Stark (Clevenger) apparatus. The condensation process was carried out in the presence of xylene until the water had been separated. An electric oven was used to dry and weigh the chitosan amide product after separating and washing it with three different solvents (methanol, hot distilled water, and ethanol) (<a href=\"#figure1\">Figure 1</a>).</p>\r\n\r\n<div id=\"figure1\">\r\n<figure class=\"image\"><img alt=\"\" height=\"145\" src=\"/media/article_images/2023/15/26/178-1653458316-Figure1.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 1.</strong> Synthesis of chitosan nanoparticles.</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Nanoparticles of chitosan and carnitine synthesis</strong><br />\r\nSynthesis of Cs-Ca NPs, by ionic gelation method, was used using TPP and Cs-Ca adduct. Cs-Ca (1 mg/ml) and TPP solution were mixed for six hours at ambient temperature with constant stirring at a ratio of 1: 2.5 (w/w %) in an acetic acid solution (1% w/v). TPP triggered ionic gelation of Cs-Ca/TPP nanoparticles. This was the first step in the process. Separation, washing, and drying of these nanoparticles resulted in the precipitate being re-suspended in water and drying [<a href=\"#r-22\">22, 23</a>].</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Characterization of L-carnitine-chitosan nanoparticles</strong><br />\r\nThese characterization tests were done in the material research laboratories at the ministry of sciences and technology environment and water research and technology director (EWRTD). Scanning Electron Microscopy (SEM) Observation<strong>–</strong>the surface morphology of N.P.s was studied using a scanning electron microscope (SEM). The Ministry of Science and Technology conducted this test as well as all other characterizations of biogenic Nano L-Carnitine. The electron microscopy-based approach’s direct observation of the nanoparticles identifies their size, shape, and surface morphology.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Animal of the study</strong><br />\r\nThe present study was conducted in the college of Veterinary medicine at the University of Basrah, in the animal house of the department of physiology. Sixty male adult albino rats (<em>Rattus rattus</em>) were used in the current study. With an average weight of 200±20g and the ages of the animals ranging from 8 to 10 weeks. They were housed for two weeks for an adaptation before the experiment. Every ten animals were housed in an individual plastic cage measured as 15x35x50cm. They were fed ad libitum with the meal of standard pellet of diet supplied by IPA (Institute for Public Accuracy). They had free access to drinking water and were kept under the exact condition of temperature (22-25) °C and light, the regime of 14 hours of light and 10 hours of darkness. The author has signed an animal welfare statement. The project was approved by the local ethical committee in the University of Al-Ameed register in number 113 on 21/2/2022.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Experimental design</strong><br />\r\nAnimals in the study were divided into six groups. Each group consists of 10 male rats used for the design of experiments as the following:<br />\r\nGroup-1 (control group): Animals received orally distilled water. Group -2 (lead acetate group): Animals received lead acetate orally at a dose of 30 mg/kg B.W. daily for 30 days. Depended on LD50, group-3 animals received lead acetate (30mg/kg B.W.) for 30 days + L-carnitine (100mg/ kg B.W) daily for 2 months. Group-4 received lead acetate (30mg/kg B.W.) for 30 days + Nano L-carnitine (100mg/ kg B.W) daily for two months. Group-5 (L-carnitine): Animals received L.C. orally at a dose of 100mg/kg B.W daily for two months. Group-6 (nano L-carnitine): Animals received LC-NPs orally at a dose of 100mg/ kg B.W daily for two months. All the experiments proceed for two months.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Sample collection</strong><br />\r\nAt the end of the treatment period, rats were dissected for sample collection. The blood sample was collected via cardiac puncture after anesthetizing the animal, according to [<a href=\"#r-24\">24</a>]. Samples were collected by using a 5ml disposable syringe. About 5ml of the collected blood was put in a non-heparinized plane tube for centrifugation at 3000 rpm for 15 minutes to obtain the serum. Then the serum was transferred to Eppendorf tubes and stored at (-4°C) until analyzed for the measurement of liver enzymes (AST, ALP, and ALT) and kidney function (creatinine and urea) to evaluate oxidative stress (MDA and GPX). Also, the brain was collected for microscopic evaluation.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Scanning electron microscopy (SEM) analysis</strong><br />\r\nThe surface morphology of N.P.s was studied using a scanning electron microscope (SEM). The Ministry of Science and Technology conducted this test as well as all other characterizations of biogenic Nano L-Carnitine. The electron microscopy-based approach’s direct observation of the nanoparticles identifies their size, shape, and surface morphology.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Biochemical parameters</strong><br />\r\nAn aspartate aminotransferase (AST) test kit is used to measure AST activity in the serum (Agape diagnostic, India code 683-562) (Clin. Chem, Acta 1976) and Dtsch. Med Wschr 1974. An alanine aminotransferase (ALT) test kit is used to measure in serum (Agappe diagnostic, India.). An alkaline phosphatase (ALP) test kit is used to measure in serum (Agappe diagnostic, India).<br />\r\nResearchers used a specialized kit (Agappe diagnostics, India) to detect the blood creatine levels. When creatinine combines with picrate in an alkaline media, a colorful complex form. A spectrophotometer that measures the absorbance at 500 nm may be used to estimate the quantity of creatinine in a specimen [<a href=\"#r-13\">13</a>]. The following reaction is used to determine urea through enzymatic means. Urea was determined according to the kit protocol of the agape diagnostic, India.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Serum malondialdehyde measurement (MDA)</strong><br />\r\nThe ability to accurately measure lipid peroxidation in disease states necessitates this method of assessing oxidative stress. MDA and 4-hydroxynonenal (4-HNE) are the natural bi-products of lipid peroxidation. One of the most commonly acknowledged methods to evaluate oxidative damage is measuring lipid peroxidation products. It is easy to use the MDA microplate assay kit to detect MDA in various samples. Thiobarbituric acid (TBA) reacts with MDA in the sample to form the MDA-TBA adduct. MDA-TBA adduct can be readily measured using a colorimeter (λ= 532 nm) [<a href=\"#r-20\">20</a>].</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Measurement of serum glutathione (GSH)</strong><br />\r\nThis kit utilizes the Sandwich-ELISA technique. The micro-ELISA plate that comes with this kit has been pre-coated with an anti-rat GPX1 antibody. Antibodies are pre-incubated in micro-ELISA plate wells before addition, and then samples or standards are added to the plate. Antibodies specific to rat GPX1 and avidin-horseradish peroxidase (HRP) conjugate are added to each micro plate well and incubated. Everything that isn’t necessary is swept away. Each well receives a substrate solution. The wells containing rat GPX1 biotinylated detection antibody and avidin-HRP conjugate will show blue.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Histological study</strong><br />\r\nThe brain was removed from each animal and fixed in 10% formalin for the preparation of a slide to investigate the histopathological changes in both the control and treated groups according to the Mescher method [<a href=\"#r-21\">21</a>] with the aid of the light microscope.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Statistical analysis</strong><br />\r\nThe Statistical Package for Social Scientists (SPSS version 18.0) and Microsoft Office Excel 2016 were used to examine the data. A one-way ANOVA with LSD post hoc test for significance was used to assess if there was a significant difference between the groups’ means. The paired t-test was used for mean comparisons, where p<0.05 values were considered significant [<a href=\"#r-25\">25</a>].</p>"
},
{
"section_number": 3,
"section_title": "RESULTS",
"body": "<p><strong>Carnitine-loaded chitosan nanoparticles (LC-NPs)</strong><br />\r\nThe nanoparticles were produced by interactions between the positively charged chitosan and the negatively charged phosphate groups of TPP in the ionic gelation technique.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Characterization of L-carnitine nanoparticles (LC-NPs)</strong><br />\r\nDifferent spectrophotometric techniques like SEM analysis investigated the morphology, elemental composition, crystalline nature, and stability of synthesized LC-NPs.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>SEM (scanning electron microscope) </strong><br />\r\nScanning electron microscope analysis of prepared chitosan nanoparticles to compare this with L-Carnitine nanoparticles (LC-NP) with different magnification pictures shows the distribution and nanoparticles size as shown in <a href=\"#figure2\">Figure 2</a>. The results of SEM images show that synthesized LC-NPs were smooth, spherical particles, singular or in aggregates with particle sizes in the range of (26.05), (33.50), and (52.10 nm)<strong>.</strong></p>\r\n\r\n<div id=\"figure2\">\r\n<figure class=\"image\"><img alt=\"\" height=\"338\" src=\"/media/article_images/2023/15/26/178-1653458316-Figure2.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 2.</strong> Scanning electron microscopy (SEM) shows the morphology and size of biogenic nanoparticles (LC-NPs).</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Liver functions</strong><br />\r\n<a href=\"#Table-1\">Table 1</a> shows the enzyme activity of AST, ALT, and ALP (alkaline phosphatase) to be significantly increased (P<0.05). While the enzyme activity of the liver (AST, ALT, and ALP) decreased significantly (P<0.05) in the nano L-carnitine and (L-carnitine) groups. Also exhibited a significant (P<0.05) decrease in lead + carnitine and lead + nano carnitine groups when compared with the lead acetate group. This indicates that the nano carnitine has more ameliorative effects on lead toxicity in animals.<br />\r\n<a href=\"#Table-2\">Table 2</a> depicts the distinguishing aspects of kidney function (creatinine and urea). When lead acetate was given, serum creatinine and urea levels rose considerably (P<0.05) compared to the other experimental groups. Nano L carnitine with lead acetate resulted in significant (P<0.05) ameliorative effects on the creatinine and urea levels in the rats. In contrast, the impact on the L-carnitine and nano L-carnitine group alone was lower than that in the control group.<br />\r\nThe results in <a href=\"#Table-3\">Table 3</a> revealed a significant (P<0.05) increase in serum MDA level in the lead acetate group (8.2 ± 0.47 nmol/ml) compared with control (5.9 ± 0.36 nmol/ml) and other treated groups. In contrast, no significant (P<0.05) difference was recorded in MDA level in L-carnitine and nano L-carnitine treated group compared with the control group. The MDA level was significantly decreased in lead acetate + nano L-carnitine treated groups (6.9 0.56) compared to the lead acetate group (8.2 0.47). This revealed a significant (P<0.05) decrease in serum GSH level in the lead acetate group (83.9 5.29) compared to the control (119.5 6.81) and other groups. In contrast, there was a substantial (P<0.05) rise in blood GSH levels in the lead acetate + nano L-carnitine, nano L-carnitine, and L-carnitine groups as compared to the lead acetate group (83.9 5.29).</p>\r\n\r\n<div id=\"Table-1\">\r\n<p><a href=\"https://jabet.bsmiab.org/table/178-1653458316-table1/\">Table-1</a><strong>Table 1.</strong> Effects of different treatments on liver enzyme activity in male rats. </p>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<div id=\"Table-2\">\r\n<p><a href=\"https://jabet.bsmiab.org/table/178-1653458316-table2/\">Table-2</a><strong>Table 2. </strong>Effects of different treatments on kidney enzyme activity in male rats.</p>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<div id=\"Table-3\">\r\n<p><a href=\"https://jabet.bsmiab.org/table/178-1653458316-table3/\">Table-3</a><strong>Table 3. </strong>Effects of different treatments on oxidative activity in male rats.</p>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Histological examination of the brain (cerebellum cortex)</strong><br />\r\nHistological examination of cerebellum sections of Control animals was found to have a normal structure, with an outside molecular layer, an inner granular layer, and a single layer of Purkinje cells in between (<a href=\"#figure3\">Figure 3A</a> and <a href=\"#figure3\">3B</a>). The treated rats in the lead acetate group exhibited signs of disrupting the three-layer structure. Degeneration, necrosis, and a reduction in the number of Purkinje cells and molecular cells were seen in the interval between Purkinje cells and a molecular granular layer (<a href=\"#figure3\">Figure 3A</a> and <a href=\"#figure3\">3B</a>). The antioxidative effect of L-carnitine was identified in the groups where the rats were administrated lead acetate with a combination of L- carnitine or nano L- carnitine. L-carnitine reduced the dangerous effect of lead, which showed normal Purkinje cells and normal arrangement of cerebellum layers (<a href=\"#figure3\">Figure 3C</a> and <a href=\"#figure3\">Figure 3D</a>). When rats were given L-carnitine and nano L-carnitine, they protected and developed brain tissue. There was no histological change in the cerebellar cortex, which revealed normal architecture and different layers of the cerebellum after treatment with l-carnitine and nano L-carnitine (<a href=\"#figure3\">Figure 3E</a> and <a href=\"#figure3\">Figure 3F</a>).</p>\r\n\r\n<div id=\"figure3\">\r\n<figure class=\"image\"><img alt=\"\" height=\"696\" src=\"/media/article_images/2023/15/26/178-1653458316-Figure3.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 3. </strong>(A) The Cerebellum section of rats in the control group (hematoxylin and Eosin .100x) showed no histopathological changes, which is composed of the outer molecular layer, inner granular layer cells, and in between, the single layer of Purkinje cells. (B) Cerebellum section of lead acetate treated rats. Shows disrupt the normal arrangement of three layers; large spaces between Purkinje’s cell layer and a molecular layer or granular layer were seen. The treated rats were administrated 30mg/kg body weight of lead acetate (hematoxylin and Eosin .400x). (C) Cerebellum Sections of Third Group (lead acetate and L-Carnitine). show normal Purkinje cells and normal arrangement of cerebellum layers. (D) Cerebellum Sections of the fourth Group (lead acetate and Nano L-Carnitine). Shows normal Purkinje cells and normal arrangement of cerebellum layers. (E) Cerebellum Sections of the fifth Group (L-Carnitine at a dose of 100mg/ kg B.W /daily for two months). Shows normal architecture, normal layers, and presence in high density central to Purkinje cells. (F) Cerebellum Sections of the sixth Group (Nano L-Carnitine at a dose of 100mg/ kg B.W /daily for two months). showed normal architecture, and normal layers of the cerebellum.</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>"
},
{
"section_number": 4,
"section_title": "DISCUSSION",
"body": "<p>Ionic gelation was used to produce nano chitosan utilizing two forms of chitosan, soluble chitosan (low molecular weight) and insoluble (high molecular weight) chitosan (high molecular weight as well as the diacylation percent) [<a href=\"#r-26\">26</a>]. Chitosan nanoparticles may be synthesized via the gelation method based on the cross-linking of anionic molecules (TPP) [<a href=\"#r-27\">27</a>]. The scanning electron microscope technique was employed to visualize the size and shape of compounds. The SEM characterizations of the synthesized chitosan nanoparticles are shown in <a href=\"#figure2\">Figure 2</a>. The image of SEM exhibited relatively smooth nano-spherical particles, singular or in aggregates with particle sizes (26.05), (33.50) and (52.10 nm). The above results agree with the authors [<a href=\"#r-28\">28</a>]. This observation of SEM images of optimized formulation (O.P.) showed that the chitosan nanoparticles were spherical. In the optimal formulation, the lecithin/chitosan nanoparticles are spherical in shape, uniform, and polydisperse, with sizes ranging from 200 to 350nm with an average size of 273nm in diameter. Saglam et al. [<a href=\"#r-29\">29</a>] mentioned that the chitosan nanoparticles loading oxaliplatin mean size and their homogeneous size distribution, without forming aggregated, besides showing they are spherical [<a href=\"#r-30\">30</a>]. Chitosan nanoparticles as a promising nanomaterial for encapsulation of pomegranate. SEM characterized their physical and structural morphology, and spherical N.P.s were successfully synthesized with a mean diameter of 174–898 nm [<a href=\"#r-31\">31</a>]. SEM characterized the morphological properties of the nanoparticles. Chitosan nanoparticles were found to have a smooth surface morphology [<a href=\"#r-32\">32</a>]. Similar results reported that SEM profiles of smooth and spherically shaped GNPs synthesized of L-asparaginase showed little homogeneity and clumps of spherical structures [<a href=\"#r-33\">33</a>].<br />\r\nIn <a href=\"#Table-1\">Table 1</a> ALT, ALP, and AST enzymes are highly concentrated in the liver. Because of liver development and repair, these enzymes are generally detected in small amounts in the bloodstream. When administrated lead acetate and compared to a control, blood liver enzymes (AST, ALT, and ALP) increased considerably; the liver was involved in detoxifying hazardous compounds such as lead acetate, which are removed by the liver following metabolism and breakdown. This process may cause cell membrane rupture, increasing serum liver enzymes. Also, rats treated with lead developed liver dysfunction and damage and histological changes demonstrated by higher ALT, AST, and ALP blood levels. L-carnitine and nano L-carnitine, when given together with lead, reduced liver damage significantly and decreased the consequences of acute toxic liver damage, according to plasma biochemical markers [<a href=\"#r-34\">34</a>].<br />\r\nThis is due to the role of the L-carnitine in increasing the production of amino acids. Especially methionine, which turns into cysteine, which is an antioxidant. It has a role in directly removing oxidative compounds or being one of the components of glutathione as an antioxidant. In addition, it protects cells from toxic substances and pollutants [<a href=\"#r-35\">35</a>]. The blood urea and creatinine levels of the control and both the nano and L-carnitine treated groups were considerably lower than the lead-induced toxicity group, as shown in <a href=\"#Table-2\">Table 2</a>. Protein metabolism’s main nitrogen-containing metabolic product is urea. Urea and creatinine levels in the blood are used as markers of renal function [<a href=\"#r-36\">36</a>]. Furthermore, a prior investigation employed elevated blood creatinine levels as a diagnostic marker of renal failure [<a href=\"#r-37\">37</a>]. The administration of both L-carnitine and Nano L-carnitine in combination with lead showed a significant reduction in blood urea and creatinine in the L-carnitine treated groups suggesting that it protects renal function against lead toxicity. Renal damage may take two forms: reversible and irreversible interstitial nephropathy. Reversible interstitial nephropathy is more frequent in long-term occupational lead exposure petering [<a href=\"#r-38\">38,39</a>].<br />\r\nThe results of oxidative stress biomarkers revealed a significant decrease in antioxidant enzyme activities (glutathione peroxidase), and an increase in the level of lipid peroxidation products (MDA) in rats given lead acetate compared to the control group (<a href=\"#Table-3\">Table 3</a>). Current results indicate a change in oxidant/antioxidant status of serum and the PbAc treated group. Moreover, the results have been attributed to oxidative stress induced by PbAc, causing ROS formation, including different pathways including hydroperoxides, singlet oxygen, and hydrogen peroxides, resulting in cellular dysfunction. These results agree with [<a href=\"#r-40\">40, 41</a>]. The extensive lipid peroxidation caused by lead exposure releases lipid hydroperoxides into the circulation, causing significant tissue oxidative damage [<a href=\"#r-42\">42,43,44</a>]. The impairment of mitochondrial oxidative phosphorylation, the collapse of mitochondrial membrane potential, the inhibition of mitochondrial respiratory enzyme activities, ATP depletion, and energy crises are only a few of the adverse outcomes seen in many models of Pb-induced toxicity [<a href=\"#r-45\">45,46</a>]. To assess lead-induced oxidative damage, researchers examined a variety of antioxidant enzymes and compounds. The most widely utilized ones are reduced, such as GSH, glutathione disulfide, GPX, superoxide dismutase (SOD), and catalase (CAT) activity [<a href=\"#r-47\">47</a>].<br />\r\nCellular molecules are protected from free radical damage by antioxidant enzymes such as CAT, SOD, and GPX [<a href=\"#r-48\">48,49</a>]. The activity of these enzymes was decreased significantly for the administration of lead acetate with L-carnitine. In contrast, these effects declined to non-significant levels compared to the control group in groups treated with nano L-carnitine and lead acetate. L-carnitine acts as an antioxidant by reducing metabolic stress [<a href=\"#r-50\">50</a>].<br />\r\nAlzheimer’s disease and geriatric depression have lately been linked to the use of L-carnitine in treating Parkinson’s disease [<a href=\"#r-51\">51,52</a>]. L-carnitine decreases exercise-induced oxidative stress, enhances antioxidant levels, and improves performance in individuals with end-stage renal failure [<a href=\"#r-53\">53,54</a>]. L-carnitine exhibited antioxidative effects by lowering MDA, hydrogen peroxide cytotoxicity, and enhancing CAT and SOD activity in hepatocytes [<a href=\"#r-55\">55</a>]. In addition to activating antioxidant enzymes like GSH, L-carnitine may also protect mitochondrial electron transport chain competence and scavenge free radical activity [<a href=\"#r-56\">56,57</a>]. Lead acetate-treated rats showed the harmful effect of lead on the brain, especially the cerebellum. The BBB is permeable to lead acetate, as evidenced by the brain accumulating lead acetate when blood Pb levels increase [<a href=\"#r-58\">58</a>]. Overwhelming epidemiological evidence shows that low-level Pb exposure causes developmental neurotoxicity [<a href=\"#r-59\">59</a>].<br />\r\nThis corresponds to the study [<a href=\"#r-60\">60</a>] on how it was assumed that lead-induced brain toxicity is primarily due to oxidative stress. There was support for this work from other researchers [<a href=\"#r-61\">61</a>]. Reduced intracellular antioxidant enzymes and lipid peroxidation products are signs of oxidative stress in the plasma and brain tissue of rats exposed to lead. Total carnitine levels in old rats were increased by giving them a daily dosage of L.C. in drinking water of 100 mg/kg of body weight [<a href=\"#r-62\">62</a>]. L-carnitine for neuroprotection in several disorders, including hypoxia-ischemia and traumatic brain injury, Alzheimer’s disease, and conditions leading to central or peripheral nervous system injury and improve energy status, decrease oxidative stress and prevent cell death in models of adult, neonatal, and pediatric brain injury [<a href=\"#r-63\">63</a>]. L-carnitine has a strong neuroprotective impact when given immediately after reperfusion from acute global cerebral ischemia [<a href=\"#r-64\">64</a>]. Carnitine is a potent free radical scavenger, which means it may protect tissues from oxidative damage by neutralizing free radicals. It also may have a role in decreasing oxidative damage reported in several hereditary neuro-metabolic illnesses. According to new studies, decreased L-carnitine concentrations have been seen with specific disorders, in part because of the compound’s interaction with accumulating toxic metabolites, particularly organic acids, or because of protein restriction [<a href=\"#r-64\">64</a>]. Because of the increased formation of reactive species in these disorders, L-carnitine supplementation may help prevent tissue shortage in this element and prevent oxidative damage. L- carnitine’s capacity to pass the blood-brain barrier suggests that it may be useful in reducing neurological damage caused by oxidative toxicity [<a href=\"#r-65\">65</a>].<br />\r\nAs L.C. may also help the body with energy metabolism and protect cells from damage, multiple studies have shown this chemical to have protective, modulatory, and trophic effects on the brain. Under metabolically challenged situations, free acetyl-CoA and ketosis may become critical for brain functioning despite the brain’s low level of -oxidation [<a href=\"#r-66\">66</a>]. L-Carnitine increased metabolic stress caused by mitochondrial dysfunction and reactive oxygen species (ROS) generation may defend cells from oxidative damage in major neurodegenerative disorders [<a href=\"#r-67\">67,68</a>]. These enzymatic antioxidants are crucial because they detoxify H<sub>2</sub>O<sub>2</sub> in the water. After chronic hypoperfusion in rats, L.C. reduced white matter damage and offered neuroprotection in old mice by enhancing the brain’s antioxidant activity [<a href=\"#r-69\">69</a>].</p>"
},
{
"section_number": 5,
"section_title": "CONCLUSIONS",
"body": "<p>The conjugated L-carnitine on chitosan nanoparticles was successfully synthesized based on the reaction between dissolving CS+LC in an aqueous acidic solution to obtain cationic chitosan+ L-carnitine and have successfully incorporated the powerful antioxidant. Biogenic L-carnitine nanoparticles (LC-NPs) alleviate the harmful effects of lead further than/ or equal to ordinary L-carnitine with a less administrated dose than L-carnitine. LC-NPs improve the liver enzyme, kidney function, antioxidant enzyme activity, and histopathological changes in the brain cerebellum returned to normal levels.</p>"
},
{
"section_number": 6,
"section_title": "ACKNOWLEDGEMENT",
"body": "<p>We would like to express our gratitude and appreciation to Dr. Ali Majeed Emara, who is invaluable in helping to do statistics, and to Dr. Labib for helping with nanomaterial work and characterization.</p>"
},
{
"section_number": 7,
"section_title": "AUTHOR CONTRIBUTIONS",
"body": "<p>All persons who meet authorship criteria are listed as authors, and all authors certify that they have participated sufficiently in the work to take public responsibility for the content, including participation in the concept, design, analysis, writing, or revision of the manuscript. Conception and design of the study: Hayder Talib Mahdi. Drafting the manuscript: Rashad Fadhil Ghadhban. Revising the manuscript: Hayder Talib Mahdi and Rashad Fadhil Ghadhban. Analysis and/or interpretation of data: Hayder Talib Mahdi and Rashad Fadhil Ghadhban.</p>"
},
{
"section_number": 8,
"section_title": "CONFLICTS OF INTEREST",
"body": "<p>There is no conflict of interest among the authors.</p>"
}
],
"figures": [
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/15/26/178-1653458316-Figure1.jpg",
"caption": "Figure 1. Synthesis of chitosan nanoparticles.",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/15/26/178-1653458316-Figure2.jpg",
"caption": "Figure 2. Scanning electron microscopy (SEM) shows the morphology and size of biogenic nanoparticles (LC-NPs).",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/15/26/178-1653458316-Figure3.jpg",
"caption": "Figure 3. (A) The Cerebellum section of rats in the control group (hematoxylin and Eosin .100x) showed no histopathological changes, which is composed of the outer molecular layer, inner granular layer cells, and in between, the single layer of Purkinje cells. (B) Cerebellum section of lead acetate treated rats. Shows disrupt the normal arrangement of three layers; large spaces between Purkinje's cell layer and a molecular layer or granular layer were seen. The treated rats were administrated 30mg/kg body weight of lead acetate (hematoxylin and Eosin .400x). (C) Cerebellum Sections of Third Group (lead acetate and L-Carnitine). show normal Purkinje cells and normal arrangement of cerebellum layers. (D) Cerebellum Sections of the fourth Group (lead acetate and Nano L-Carnitine). Shows normal Purkinje cells and normal arrangement of cerebellum layers. (E) Cerebellum Sections of the fifth Group (L-Carnitine at a dose of 100mg/ kg B.W /daily for two months). Shows normal architecture, normal layers, and presence in high density central to Purkinje cells. (F) Cerebellum Sections of the sixth Group (Nano L-Carnitine at a dose of 100mg/ kg B.W /daily for two months). showed normal architecture, and normal layers of the cerebellum.",
"featured": false
}
],
"authors": [
{
"id": 476,
"affiliation": [
{
"affiliation": "Department of Physiology, Pharmacology and Biochemistry, College of Veterinary Medicine, University of Basrah, Basrah, Iraq"
}
],
"first_name": "Hayder Talib",
"family_name": "Mahdi",
"email": "hayder@alameed.edu.iq",
"author_order": 1,
"ORCID": null,
"corresponding": true,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "Hayder Talib Mahdi, PhD; Department of Physiology, Pharmacology and Biochemistry, College of Veterinary Medicine, University of Basrah, Basrah, Iraq, e-mail: hayder@alameed.edu.iq",
"article": 118
},
{
"id": 477,
"affiliation": [
{
"affiliation": "Department of Physiology, college of medicine, University of Al-Ameed, Karbala, Iraq"
}
],
"first_name": "Rashad Fadhil",
"family_name": "Ghadhban",
"email": null,
"author_order": 2,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 118
}
],
"views": 1699,
"downloads": 207,
"references": [
{
"id": 3729,
"serial_number": 1,
"pmc": null,
"reference": "Abdulidha NA, Jaccob AA, AL-Moziel MS. Protective effects of Co-Q10, Ginkgo biloba, l-carnitine on brain, kidney, liver, and endocrine system against sub-acute heavy metals toxicity in male rats. Toxicology and Environmental Health Sciences. 2020; 12(4): 331-341. DOI: 10.1007/s13530-020-00061-7.",
"DOI": null,
"article": 118
},
{
"id": 3730,
"serial_number": 2,
"pmc": null,
"reference": "Alwan S, Al-Saeed M, Abid H. Safety assessment and biochemical evaluation of biogenic silver nanoparticles (using bark extract of C. zeylanicum) in Rattus norvegicus rats. Baghdad Journal of Biochemistry and Applied Biological Sciences. 2021; 2(03): 138-150. DOI: org/10.47419/bjbabs.v2i03.67.",
"DOI": null,
"article": 118
},
{
"id": 3731,
"serial_number": 3,
"pmc": null,
"reference": "Alwan SH, Al-Saeed MH. Biosynthesized silver nanoparticles (using Cinnamomum zeylanicum bark extract) improve the fertility status of rats with polycystic ovarian syndrome. Biocatalysis and Agricultural Biotechnology. 2021; 38(3) DOI: org/10.1016/j.bcab.2021.102217.",
"DOI": null,
"article": 118
},
{
"id": 3732,
"serial_number": 4,
"pmc": null,
"reference": "Nam SM, Seo JS, Nah, SS, Chang, BJ. Effects of ascorbic acid on osteopontin expression and axonal myelination in the developing cerebellum of lead-exposed rat pups. International journal of environmental research and public health. 2019; 16(6): 983. DOI: org/10.3390/ijerph16060983.",
"DOI": null,
"article": 118
},
{
"id": 3733,
"serial_number": 5,
"pmc": null,
"reference": "Zaboon MH, Saleh AA, Al-Lami HS. Synthesis, characterization and cytotoxicity investigation of chitosan-amino acid derivatives nanoparticles in human breast cancer cell lines. Journal of the Mexican Chemical Society. 2021; 65(2): 178-188. DOI: org/10.29356/jmcs.v65i2.1265.",
"DOI": null,
"article": 118
},
{
"id": 3734,
"serial_number": 6,
"pmc": null,
"reference": "Abdul HM, Butterfield DA. Involvement of PI3K/PKG/ERK1/2 signaling pathways in cortical neurons to trigger protection by cotreatment of acetyl-L-carnitine and alpha-lipoic acid against HNE-mediated oxidative stress and neurotoxicity: implications for Alzheimer’s disease. Free Radical Biology and Medicine. 2007; 42(3): 371–384. DOI: 10.1016/j. freeradbiomed.2006.11.006.",
"DOI": null,
"article": 118
},
{
"id": 3735,
"serial_number": 7,
"pmc": null,
"reference": "Ahmed HA, Ali HA, Mutar TF. Protective effects of olive leaf extract against reproductive toxicity of the lead acetate in rats. Environmental Science and Pollution Research. 2021; 28(44): 63102-63110. DOI: 10.1007/s11356-021-15240-3.",
"DOI": null,
"article": 118
},
{
"id": 3736,
"serial_number": 8,
"pmc": null,
"reference": "Ahmed TA, Aljaeid BM. Preparation, characterisation, and potential application of chitosan, chitosan derivatives, and chitosan metal nanoparticles in pharmaceutical drug delivery. Drug design, development and therapy. 2016; 10(483): DOI: 10.2147/DDDT.S99651.",
"DOI": null,
"article": 118
},
{
"id": 3737,
"serial_number": 9,
"pmc": null,
"reference": "Akbarian A, Michiels J, Degroote J, Majdeddin M, Golian A, De Smet S. Association between heat stress and oxidative stress in poultry; mitochondrial dysfunction and dietary interventions with phytochemicals. Journal of animal science and biotechnology. 2016; 7(1): 1-14. DOI: org/10.1186/s40104-016-0097-5.",
"DOI": null,
"article": 118
},
{
"id": 3738,
"serial_number": 10,
"pmc": null,
"reference": "Al-Dulimi AG, Al-Saffar AZ, Sulaiman GM, Khalil KA, Khashan KS, Al-Shmgani HS, et al. Immobilisation of l-asparaginase on gold nanoparticles for novel drug delivery approach as anticancer agent against human breast carcinoma cells. Journal of Materials Research and Technology. 2020; 9(6): 15394-15411. DOI: org/10.1016/j.jmrt.2020.10.021.",
"DOI": null,
"article": 118
},
{
"id": 3739,
"serial_number": 11,
"pmc": null,
"reference": "Al-Fartosy AJM, Awad NA, Alsalimi SA. Insulin resistance and specific biomarkers in blood and urine of type 2 diabetic patients with or without nephropathy in Basrah, Iraq. African Journal of Biochemistry Research. 2020; 14(4):125-134. DOI: 10.5897// AJBR2020.1101.",
"DOI": null,
"article": 118
},
{
"id": 3740,
"serial_number": 12,
"pmc": null,
"reference": "Alharthi WA, Hamza RZ, Elmahdi MM, Abuelzahab HS, Saleh H. Selenium and L-carnitine ameliorate reproductive toxicity induced by cadmium in male mice. Biological Trace Element Research. 2020; 197(2): 619-627. DOI: 10.1007/s12011-019-02016-7.",
"DOI": null,
"article": 118
},
{
"id": 3741,
"serial_number": 13,
"pmc": null,
"reference": "Ali R, Hou GJ, Zhu ZG, Yan QB, Zheng QR, Su G. Stable mixed group II (Ca, Sr) and XIV (Ge, Sn) lead-free perovskite solar cells. Journal of Materials Chemistry A. 2018; 6(19): 9220-9227. DOI: org/10.1039/C8TA01490F.",
"DOI": null,
"article": 118
},
{
"id": 3742,
"serial_number": 14,
"pmc": null,
"reference": "Al-Okaily BN, Murad HF. Role of alpha lipoic acid in protecting testes of adult rats from lead toxicity. Iraqi Journal of Veterinary Sciences. 2021; 35(2): 305-312. DOI: 10.33899/ijvs.2020.126814.1386.",
"DOI": null,
"article": 118
},
{
"id": 3743,
"serial_number": 15,
"pmc": null,
"reference": "Anjum MR, Reddy PS. Recovery of lead‐induced suppressed reproduction in male rats by testosterone. Andrologia. 2015; 47(5): 560-567. DOI: 10.1111/and.12303.",
"DOI": null,
"article": 118
},
{
"id": 3744,
"serial_number": 16,
"pmc": null,
"reference": "Assi MA, Hezmee MN, Sabri MY, Rajion MA. The detrimental effects of lead on human and animal health. Veterinary world. 2016; 9(6): 660-671. DOI: 10.14202/vetworld. 2016.660-671.",
"DOI": null,
"article": 118
},
{
"id": 3745,
"serial_number": 17,
"pmc": null,
"reference": "Al-Megrin WA, Alkhuriji AF, Yousef AO, Habotta OA, Kassab RB, Metwally D, et al. Antagonistic efficacy of luteolin against lead acetate exposure-associated with hepatotoxicity is mediated via antioxidant, anti-inflammatory, and anti-apoptotic activities. Antioxidants. 2019; 9(1): 10. DOI: 10.3390/antiox9010010.",
"DOI": null,
"article": 118
},
{
"id": 3746,
"serial_number": 18,
"pmc": null,
"reference": "Basiouny FES, Mohamed FB, Ehab T. Ameliorating role of L-carnitine and Ginkgo biloba extract on pentylenetetrazole induced bone marrow injury in epileptic seizures disease in the rat. GSC Advanced Research and Reviews. 2020; 5(2): 001-011. DOI: 10.30574/gscarr.2020.5.2.0092.",
"DOI": null,
"article": 118
},
{
"id": 3747,
"serial_number": 19,
"pmc": null,
"reference": "Caito SW, Aschner M. Mitochondrial redox dysfunction and environmental exposures. Antioxidants & redox signaling. 2015; 23(6): 578-595. DOI: 10.1089/ars.2015.6289.",
"DOI": null,
"article": 118
},
{
"id": 3748,
"serial_number": 20,
"pmc": null,
"reference": "Debnath SK, Saisivam S, Debanth M, Omri A. Development and evaluation of Chitosan nanoparticles based dry powder inhalation formulations of Prothionamide. PloS one. 2018; 13(1): e0190976. DOI: 10.1371/journal. pone.0190976.",
"DOI": null,
"article": 118
},
{
"id": 3749,
"serial_number": 21,
"pmc": null,
"reference": "Dkhil MA, Moneim AEA, Al-Quraishy S. Indigofera oblongifolia ameliorates lead acetate-induced testicular oxidative damage and apoptosis in a rat model. Biological trace element research. 2016; 173(2): 354-361. DOI: 10.1007/s12011-016-0689-0.",
"DOI": null,
"article": 118
},
{
"id": 3750,
"serial_number": 22,
"pmc": null,
"reference": "Dugard P. Randomisation tests: A new gold standard. Journal of Contextual Behavioral Science. 2014; 3(1): 65-68. DOI: org/10.1016/j.jcbs.2013.10.001.",
"DOI": null,
"article": 118
},
{
"id": 3751,
"serial_number": 23,
"pmc": null,
"reference": "El‐Ghaffar MA, Hashem MS. Immobilisation of α‐amylase onto chitosan and its amino acid condensation adducts. Journal of applied polymer science. 2009; 112(2): 805-814. DOI: org/10.1002/app.29292.",
"DOI": null,
"article": 118
},
{
"id": 3752,
"serial_number": 24,
"pmc": null,
"reference": "El‐Ghaffar MA, Atia KS, Hashem MS. Synthesis and characterisation of binary copolymers of methyl methacrylate with glycidyl methacrylate and 2‐hydroxy ethyl methacrylate as carriers for cellulase. Journal of Applied Polymer Science. 2010; 117(2): 629-638. DOI: org/10.1002/app.30063.",
"DOI": null,
"article": 118
},
{
"id": 3753,
"serial_number": 25,
"pmc": null,
"reference": "Essawy AE, El-Sayed SA, Tousson E, El-gawad A, Horeya S, Alhasani RH, et al. Anti-kindling effect of Ginkgo biloba leaf extract and L-carnitine in the pentylenetetrazol model of epilepsy. Environmental Science and Pollution Research. 2022; 1-15. DOI: 10.1007/s11356-022-19251-6.",
"DOI": null,
"article": 118
},
{
"id": 3754,
"serial_number": 26,
"pmc": null,
"reference": "Fajardo AR, Pereira AG, Martins AF, Paulino AT, Muniz EC, Hsieh YL. Chitin and chitosan-based (NANO) composites. John Wiley & Sons Inc. 2017; 671-700: DOI: 10.1002/9781119441632 . ch147.",
"DOI": null,
"article": 118
},
{
"id": 3755,
"serial_number": 27,
"pmc": null,
"reference": "Fatouros IG, Douroudos I, Panagoutsos S, Pasadakis P, Nikolaidis MG, Chatzinikolaou,A, et al. Effects of L-carnitine on oxidative stress responses in patients with renal disease. Med Sci Sports Exerc. 2010; 42(10): 1809-18. DOI: 10.1249/MSS.0b013e3181dbacab.",
"DOI": null,
"article": 118
},
{
"id": 3756,
"serial_number": 28,
"pmc": null,
"reference": "Ferreira GC, McKenna MC. L-Carnitine and acetyl-L-carnitine roles and neuroprotection in developing brain. Neurochemical research. 2017; 42(6): 1661-1675. DOI: 10.1007/s11064-017-2288-7.",
"DOI": null,
"article": 118
},
{
"id": 3757,
"serial_number": 29,
"pmc": null,
"reference": "Glicklich D, Frishman WH. The Case for Cadmium and Lead Heavy Metal Screening. The American journal of the medical sciences. 2021; 362(4): 344-354. DOI: 10.1016/j.amjms.2021.05.019.",
"DOI": null,
"article": 118
},
{
"id": 3758,
"serial_number": 30,
"pmc": null,
"reference": "Hall JA, Yerramilli M, Obare E, Yerramilli M, Jewell DE. Comparison of serum concentrations of symmetric dimethylarginine and creatinine as kidney function biomarkers in cats with chronic kidney disease. Journal of veterinary internal medicine. 2014; 28(6): 1676-1683. DOI: 10.1111/jvim.12445.",
"DOI": null,
"article": 118
},
{
"id": 3759,
"serial_number": 31,
"pmc": null,
"reference": "Hammed MS. Evaluation of Performance of Date Palm Pollen on Urea and Creati-nine Levels in Adult Female Rats Exposed to Lead Acetate Intoxication. International Journal of Biomedicine and Advanced Researches. 2015; 6(1) 20: DOI: org/10.7439/ijbar.v6i1.1565.",
"DOI": null,
"article": 118
},
{
"id": 3760,
"serial_number": 32,
"pmc": null,
"reference": "Hinerfeld D, Traini MD, Weinberger RP. Cochran B, Doctrow SR, Harry J, et al. Endogenous mitochondrial oxidative stress: neurodegeneration, proteomic analysis, specific respiratory chain defects, and efficacious antioxidant therapy in superoxide dismutase two null mice. Journal of Neurochemical. 2004; 88(3): 657–667. DOI: 10.1046/j.1471-4159.2003.02195.x.",
"DOI": null,
"article": 118
},
{
"id": 3761,
"serial_number": 33,
"pmc": null,
"reference": "Hou P, Wang F, Luo B, Li A, Wang C, Shabala L, et al. Antioxidant enzymatic activity and osmotic adjustment as components of the drought tolerance mechanism in Carex duriuscula. Plants. 2021; 10(3): 436. DOI: 10.3390/plants10030436.",
"DOI": null,
"article": 118
},
{
"id": 3762,
"serial_number": 34,
"pmc": null,
"reference": "Hudson SA, Tabet N. Acetyl‐l‐carnitine for dementia. Cochrane Database of Systematic Reviews. 2003; (2): DOI: 10.1002/14651858.CD003158.",
"DOI": null,
"article": 118
},
{
"id": 3763,
"serial_number": 35,
"pmc": null,
"reference": "Ilk S, Sağlam N, Özgen M, Korkusuz F. Chitosan nanoparticles enhances the anti-quorum sensing activity of kaempferol. International journal of biological macromolecules. 2017; 94: 653-662. DOI: 10.1016/j.ijbiomac.2016.10.068.",
"DOI": null,
"article": 118
},
{
"id": 3764,
"serial_number": 36,
"pmc": null,
"reference": "ackie T, Haleagrahara N, Chakravarthi S. Antioxidant effects of Etlingera elatior flower extract against lead acetate-induced perturbations in free radical scavenging enzymes and lipid peroxidation in rats. BMC research notes. 2011; 4(1): 1-8. DOI: 10.1186/1756-0500-4-67.",
"DOI": null,
"article": 118
},
{
"id": 3765,
"serial_number": 37,
"pmc": null,
"reference": "Juliet PA, Joyee AG, Jayaraman G, Mohankumar MN, Panneerselvam C. Effect of L-carnitine on nucleic acid status of aged rat brain. Experimental neurology. 2005; 191(1): 33-40. DOI: 10.1016/j.expneurol. 2004.09.009.",
"DOI": null,
"article": 118
},
{
"id": 3766,
"serial_number": 38,
"pmc": null,
"reference": "Khatab LA, Abdel-Raheem IT, Ghoneim AI. Protective effects of melatonin and l-carnitine against methotrexate-induced toxicity in isolated rat hepatocytes. Naunyn-Schmiedeberg’s Archives of Pharmacology. 2022; 395(1): 87-97. DOI: 10.1007/s00210-021-02176-1.",
"DOI": null,
"article": 118
},
{
"id": 3767,
"serial_number": 39,
"pmc": null,
"reference": "Kumar A, Kumar A, MMS CP, Chaturvedi AK, Shabnam AA, Subrahmanyam G, et al. Lead toxicity: health hazards, influence on food chain, and sustainable remediation approaches. International journal of environmental research and public health. 2020; 17(7): 2179. DOI: 10.3390/ijerph17072179.",
"DOI": null,
"article": 118
},
{
"id": 3768,
"serial_number": 40,
"pmc": null,
"reference": "Kumar SD and Kumar SS. Effect of heat treatment conditions on ballistic behaviour of various zones of friction stir welded magnesium alloy joints. Transactions of Nonferrous Metals Society of China. 2021; 31(1): 156-166. DOI: org/10.1016/S1003-6326(20)65484-X.",
"DOI": null,
"article": 118
},
{
"id": 3769,
"serial_number": 41,
"pmc": null,
"reference": "Li JL, Wang QY, Luan HY, Kang ZC, Wang CB. Effects of L-carnitine against oxidative stress in human hepatocytes: involvement of peroxisome proliferator-activated receptor alpha. Journal of biomedical science. 2012; 19(1): 1-9. DOI: org/10.1186/1423-0127-19-32.",
"DOI": null,
"article": 118
},
{
"id": 3770,
"serial_number": 42,
"pmc": null,
"reference": "Lopes ACBA, Peixe TS, Mesas AE, Paoliello M. Lead exposure and oxidative stress: a systematic review. Reviews of Environmental Contamination and Toxicology. 2016; 236: 193-238. DOI: 10.1007/978-3-319-20013-2_3.",
"DOI": null,
"article": 118
},
{
"id": 3771,
"serial_number": 43,
"pmc": null,
"reference": "Matos BN, Pereira MN, Bravo MDO, Cunha-Filho M, Saldanha-Araújo F, Gratieri T, et al. Chitosan nanoparticles loading oxaliplatin as a mucoadhesive topical treatment of oral tumors: Iontophoresis further enhances drug delivery ex vivo. International journal of biological macromolecules. 2020; 154: 1265-1275. DOI: 10.1016/j.ijbiomac.2019.11.001.",
"DOI": null,
"article": 118
},
{
"id": 3772,
"serial_number": 44,
"pmc": null,
"reference": "Morris GA, Kök SM, Harding SE, Adams GG. Polysaccharide drug delivery systems based on pectin and chitosan. Biotechnology and genetic engineering reviews. 2010; 27(1): 257-284. DOI: 10.1080/02648725.2010.10648153.",
"DOI": null,
"article": 118
},
{
"id": 3814,
"serial_number": 45,
"pmc": null,
"reference": "Munir M, Habib M, Khan SA, Kim MH, Lee S, Song TK, et al. Energy storage and piezoelectric properties of lead‐free SrTiO3-modified 0.965 Bi0. 5Na0. 5TiO3–0.035 BaTiO3 ceramics. Journal of Materials Science: Materials in Electronics. 2021; 32(8): 10712-10725. DOI: org/10.1007/s10854-021-05728-6.",
"DOI": null,
"article": 118
},
{
"id": 3815,
"serial_number": 46,
"pmc": null,
"reference": "Abd El-Hack ME, El-Saadony MT, Shafi ME, Zabermawi NM, Arif M, Batiha GE, et al. Antimicrobial and antioxidant properties of chitosan and its derivatives and their applications: A review. International Journal of Biological Macromolecules. 2020; 164: 2726-2744. DOI: org/10.1016/j.ijbiomac.2020.08.153.",
"DOI": null,
"article": 118
},
{
"id": 3816,
"serial_number": 47,
"pmc": null,
"reference": "Ommati MM, Jamshidzadeh A, Heidari R, Sun Z, Zamiri MJ, Khodaei F, et al. Carnosine and histidine supplementation blunt lead-induced reproductive toxicity through antioxidative and mitochondria-dependent mechanisms. Biological trace element research. 2019; 187(1): 151-162. DOI: 10.1007/s12011-018-1358-2.",
"DOI": null,
"article": 118
},
{
"id": 3817,
"serial_number": 48,
"pmc": null,
"reference": "Owen L, Sunram-Lea SI. Metabolic agents that enhance ATP can improve cognitive functioning: a review of the evidence for glucose, oxygen, pyruvate, creatine, and L-carnitine. Nutrients. 2011; 3(8): 735-755. DOI: 10.3390/nu3080735.",
"DOI": null,
"article": 118
},
{
"id": 3818,
"serial_number": 49,
"pmc": null,
"reference": "Ozsoy SY, Ozsoy B, Ozyildiz Z, Aytekin I. Protective effect of L-carnitine on experimental lead toxicity in rats: a clinical, histopathological and immunohistochemical study. Biotechnic & Histochemistry. 2011; 86(6): 436-443. DOI: 10.3109/10520295.2010.529825.",
"DOI": null,
"article": 118
},
{
"id": 3819,
"serial_number": 50,
"pmc": null,
"reference": "Patwa J, Sharma A, Flora SJ. Arsenic, cadmium, and lead. In Reproductive and Developmental Toxicology. 2022; (pp. 547-571). DOI: 10.1016/B978-0-12-804239-7.00031-7.",
"DOI": null,
"article": 118
},
{
"id": 3820,
"serial_number": 51,
"pmc": null,
"reference": "Pettegrew JW, Levine J, McClure RJ. Acetyl-L-carnitine physical-chemical, metabolic, and therapeutic properties: relevance for its mode of action in Alzheimer’s disease and geriatric depression. Molecular psychiatry. 2000; 5(6): 616-632. DOI: 10.1038/sj.mp.4000805.",
"DOI": null,
"article": 118
},
{
"id": 3821,
"serial_number": 52,
"pmc": null,
"reference": "Qian Y, Wang L, Evelyn T. Tiffany-Castiglioni. Lead-Modulated Gene Expression in the Central Nervous System. Reference Module in Biomedical Sciences. 2014; DOI: 10.1016/B978-0-12-801238-3.01926-7.",
"DOI": null,
"article": 118
},
{
"id": 3822,
"serial_number": 53,
"pmc": null,
"reference": "Qiao K, Xu L, Tang J, Wang Q, Lim KS, Hooper G, et al. The advances in nanomedicine for bone and cartilage repair. Journal of Nanobiotechnology. 2022; 20(1): 1-42. DOI: org/10.1186/s12951-022-01342-8.",
"DOI": null,
"article": 118
},
{
"id": 3823,
"serial_number": 54,
"pmc": null,
"reference": "Rahman Z, Singh VP. The relative impact of toxic heavy metals (THMs) (arsenic (As), cadmium (Cd), chromium (Cr)(VI), mercury (Hg), and lead (Pb)) on the total environment: an overview. Environmental monitoring and assessment. 2019; 191(7): 1-21. DOI: 10.1007/s10661-019-7528-7.",
"DOI": null,
"article": 118
},
{
"id": 3824,
"serial_number": 55,
"pmc": null,
"reference": "Rathbone MJ, Hadgraft J, Roberts MS. Modified-release drug delivery technology. Drugs and the pharmaceutical sciences. 2003; 126: 101-114. DOI: org/10.1201/9780203910337.",
"DOI": null,
"article": 118
},
{
"id": 3825,
"serial_number": 56,
"pmc": null,
"reference": "Renugadevi J, Prabu SM. Quercetin protects against oxidative stress-related renal dysfunction by cadmium in rats. Experimental and Toxicologic Pathology. 2010; 62(5): 471-481. DOI: 10.1016/j.etp.2009.06.006.",
"DOI": null,
"article": 118
},
{
"id": 3826,
"serial_number": 57,
"pmc": null,
"reference": "Ribas GS, Vargas CR, Wajner M. L-carnitine supplementation as a potential antioxidant therapy for inherited neurometabolic disorders. Gene. 2014; 533(2): 469-476. DOI: 10.1016/j.gene.2013.10.017.",
"DOI": null,
"article": 118
},
{
"id": 3827,
"serial_number": 58,
"pmc": null,
"reference": "Rosenthal RE, Williams R, Bogaert YE, et al. Prevention of postischemic canine neurological injury through potentiation of brain energy metabolism by acetyl-L-carnitine. Stroke. 1992; 23(9): 1312-1317. DOI: 10.1161/01.str.23.9.1312.",
"DOI": null,
"article": 118
},
{
"id": 3828,
"serial_number": 59,
"pmc": null,
"reference": "Al-Mosawi RAS, Al-Fartosi KG. Oxidant and Antioxidant Status of patients with Sickle cell–ß Thalassemia in Thi-Qar Province, Iraq. Research Journal of Pharmacy and Technology. 2020; 13(10): 4796-4800. DOI: 10.5958/0974-360X.2020.00843.4.",
"DOI": null,
"article": 118
},
{
"id": 3829,
"serial_number": 60,
"pmc": null,
"reference": "Salama A, Elgohary R. L-carnitine and Co Q10 ameliorate potassium dichromate-induced acute brain injury in rats targeting AMPK/AKT/NF-κβ. International Immunopharmacology. 2021; 101: 107867. DOI: 10.1016/j.intimp. 2021.107867.",
"DOI": null,
"article": 118
},
{
"id": 3830,
"serial_number": 61,
"pmc": null,
"reference": "Sawicka AK, Renzi G, Olek RA. The bright and the dark sides of L-carnitine supplementation: a systematic review. Journal of the International Society of Sports Nutrition. 2020; 17(1): 1-10. DOI: 10.1186/s12970-020-00377-2.",
"DOI": null,
"article": 118
},
{
"id": 3831,
"serial_number": 62,
"pmc": null,
"reference": "Shaban NZ, El-Kader A, Sara E, Mogahed FA, El-Kersh, MA, Habashy NH. Synergistic protective effect of Beta vulgaris with meso-2, 3-dimercaptosuccinic acid against lead-induced neurotoxicity in male rats. Scientific Reports. 2021; 11(1): 1-18. DOI: org/10.1038/s41598-020-80669-4.",
"DOI": null,
"article": 118
},
{
"id": 3832,
"serial_number": 63,
"pmc": null,
"reference": "Soltanzadeh M, Peighambardoust SH, Ghanbarzadeh B, et al. Chitosan nanoparticles as a promising nanomaterial for encapsulation of pomegranate (Punica granatum L.) peel extract as a natural source of antioxidants. Nanomaterials. 2021; 11(6): 1439. DOI: org/10.3390/nano11061439.",
"DOI": null,
"article": 118
},
{
"id": 3833,
"serial_number": 64,
"pmc": null,
"reference": "Sudjarwo SA, Giftania Wardani K. Protective effect of curcumin on lead acetate-induced testicular toxicity in Wistar rats. Research in pharmaceutical sciences. 2017; 12(5): 381. DOI: 10.4103/1735-5362.213983.",
"DOI": null,
"article": 118
},
{
"id": 3834,
"serial_number": 65,
"pmc": null,
"reference": "Surai PF. Carnitine Enigma: From Antioxidant Action to Vitagene Regulation Part 2. Transcription Factors and Practical Applications. J Veter Sci Med. 2015; 3(2): 17. DOI: 10.13188/2325-4645.1000017.",
"DOI": null,
"article": 118
},
{
"id": 3835,
"serial_number": 66,
"pmc": null,
"reference": "anaka Y, Sasaki R, Fukui F, Waki H, Kawabata T, Okazaki M, et al. Acetyl-L-carnitine supplementation restores decreased tissue carnitine levels and impaired lipid metabolism in aged rats. Journal of Lipid Research. 2004; 45: 729–735. DOI: 10.1194/jlr.M300425-JLR200.",
"DOI": null,
"article": 118
},
{
"id": 3836,
"serial_number": 67,
"pmc": null,
"reference": "Tang ZX, Qian JQ, Shi LE. Preparation of chitosan nanoparticles as carrier for immobilised enzyme. Applied Biochemistry and Biotechnology. 2007; 136(1): 77-96. DOI: 10.1007/BF02685940.",
"DOI": null,
"article": 118
},
{
"id": 3837,
"serial_number": 68,
"pmc": null,
"reference": "Taskin T, Dogan M, Arabaci T. Bioassay-guided isolation and antiproliferative efficacy of extract loaded in chitosan nanoparticles and LC-QTOF-MS/MS analysis of Achillea magnifica. South African Journal of Botany. 2020; 133: 236-244. DOI: org/10.1016/j.sajb.2020.08.002.",
"DOI": null,
"article": 118
},
{
"id": 3838,
"serial_number": 69,
"pmc": null,
"reference": "Tiwari S, Kumar V, Randhawa S, Verma SK. Preparation and characterisation of extracellular vesicles. American Journal of Reproductive Immunology. 2021; 85(2): e13367. DOI: 10.1111/aji.13367.",
"DOI": null,
"article": 118
},
{
"id": 3839,
"serial_number": 70,
"pmc": null,
"reference": "Rogers WA. The tangled web of medical and commercial interests. Health Expectations: an International Journal of Public Participation in Health Care and Health Policy. 2007; 10(1): 1. DOI: 10.1111/j.1369-7625.2007.00432.x.",
"DOI": null,
"article": 118
},
{
"id": 3840,
"serial_number": 71,
"pmc": null,
"reference": "Ueno Y, Saito A, Nakata J, Kamagata K, Taniguchi D, Motoi Y, et al. Possible Neuroprotective Effects of l-Carnitine on White-Matter Microstructural Damage and Cognitive Decline in Hemodialysis Patients. Nutrients. 2021; 13(4): 1292. DOI: 10.3390/nu13041292.",
"DOI": null,
"article": 118
},
{
"id": 3841,
"serial_number": 72,
"pmc": null,
"reference": "Uwikor FK, Nwachuku EO, Igwe F, Echonwere B, Bartimaeus ES. Evaluation of Haematological Changes In Lead-Acetate-Induced Albino Rats Treated With Aqueous Extract Of Hypoestes Rosea Leaf. European Journal of Biomedical. 2020; 7(2): 509-517. DOI: 10.9734/jocamr/2020/ v9i130135.",
"DOI": null,
"article": 118
},
{
"id": 3842,
"serial_number": 73,
"pmc": null,
"reference": "Virgolini MB and Aschner M. Molecular mechanisms of lead neurotoxicity. In Advances in neurotoxicology. 2021; 5: 159-213 Academic Press. DOI: 10.1016/bs.ant.2020.11.002.",
"DOI": null,
"article": 118
},
{
"id": 3843,
"serial_number": 74,
"pmc": null,
"reference": "Virmani A and Binienda Z. Role of carnitine esters in brain neuropathology. Mol. Aspects Med. 2004; 25(6): 533–549. DOI: org/10.1016/j.mam.2004.06.003.",
"DOI": null,
"article": 118
},
{
"id": 3844,
"serial_number": 75,
"pmc": null,
"reference": "Virmani MA and Maria C. The Role of l-Carnitine in Mitochondria, Prevention of Metabolic Inflexibility and Disease Initiation. International Journal of Molecular Sciences. 2022; 23(5): 2717. DOI: 10.3390/ijms23052717.",
"DOI": null,
"article": 118
},
{
"id": 3845,
"serial_number": 76,
"pmc": null,
"reference": "Yang L, Li X, Jiang A, Li X, Chang W, Chen J, et al. Metformin alleviates lead-induced mitochondrial fragmentation via AMPK/Nrf2 activation in SH-SY5Y cells. Redox biology. 2020; 36: 101626. DOI: org/10.1016/j.redox.2020.101626.",
"DOI": null,
"article": 118
}
]
},
{
"id": 119,
"slug": "178-1652598688-codon-usage-bias-and-purifying-selection-identified-in-cirrhinus-reba-mitogenome",
"featured": false,
"slider": false,
"issue": "Vol5 Issue3",
"type": "original_article",
"manuscript_id": "178-1652598688",
"recieved": "2022-05-15",
"revised": null,
"accepted": "2022-06-28",
"published": "2022-07-07",
"pdf_file": "https://jabet.bsmiab.org/media/pdf_file/2023/12/178-1652598688.pdf",
"title": "Codon usage bias and purifying selection identified in Cirrhinus reba mitogenome",
"abstract": "<p>In our previous study, we described sequence assembly and organization of the complete mitochondrial genome of a threatened labeonine fish, <em>Cirrhinus reba</em> (GenBank accession no.: MN862482). In this study, our attempts were to find out any mutation or selection pressures and codon usage patterns existing in the mitogenome of the same fish. We applied bioinformatics tools to measure important gene parameters including AT/GC skewness, codon adaptation index (CAI), the effective number of codons (ENc) and GC percentages of each protein coding gene. We found an overrepresentation of A and C resulting a lower number of T and G bases, respectively, where AT-skew was slightly positive, and GC-skew was slightly negative. Except for ND6, all protein coding genes (PCGs) had negative GC-skew, which indicated the higher occurrence of Cs. Based on comparative selective strength analysis with the PCGs of two related mitogenomes, we revealed that most of the PCGs (except for ND1, COX2 and ND5 genes) retained the dN/dS or Ka/Ks values less than 0.1 implying that they evolved under strong purifying selection. We further analyzed the codon frequency and relative synonymous codon usage (RSCU) and observed a total of 3802 codons which were used for coding 20 amino acids by a standard set of 64 codons. The amino acids Leucine and Serine were encoded each by six different codons, whereas rest of the amino acids was encoded by either two or four codons. We identified a total of 25 RSCU values (> 1) and revealed 12 codons as “overpresented” that implied for codon usage bias to engage in highly expressed genes for efficient protein synthesis via translational selection. The existence of codon usage biasness rolling in translational selection and the signs of purifying selection identified in PCGs suggest obvious conservation of this threatened fish species.</p>",
"journal_reference": "J Adv Biotechnol Exp Ther. 2022; 5(3): 605-614.",
"academic_editor": "Md. Masudur Rahman, PhD; Sylhet Agricultural University, Bangladesh",
"cite_info": "Islam MN, Sultana S. Codon usage bias and purifying selection identified in Cirrhinus reba mitogenome. J Adv Biotechnol Exp Ther. 2022; 5(3): 605-614.",
"keywords": [
"RSCU analysis",
"Selective pressure",
"Negative selection",
"Evolution rates",
"Codon usage",
"Mitochondrial genes"
],
"DOI": "10.5455/jabet.2022.d139",
"sections": [
{
"section_number": 1,
"section_title": "INTRODUCTION",
"body": "<p>Mutation rate is high in mitochondrial genomes because of the free radicals which are generated by the respiration inside the mitochondria and remain very close proximity to the mitochondrial DNA. Consequently, mitochondrial genes experience much higher and faster evolution rates than nuclear genes. In the mitogenomes of different species, mutation pressure creates variation in the frequencies of the nucleotide bases at both synonymous and nonsynonymous sites [<a href=\"#r-1\">1-2</a>]. Due to the asymmetric type of replication of two strands of mitochondrial genome, the mutation rates are not equal which means that the frequencies of G and C are not same, and the frequencies of A and U are also different [<a href=\"#r-1\">1-3</a>].<br />\r\nAlthough mitochondrial gene order and composition are highly conserved, nucleotide sequences show rapid divergence [<a href=\"#r-4\">4</a>]. The famous population geneticist, Motoo Kimura postulated the neutral theory of molecular evolution, claiming that in most instances where a genetic change spreads across a population caused by genetic drift acting on neutral mutations [<a href=\"#r-5\">5</a>]. However, the change is not necessarily reflected in phenotypic change [<a href=\"#r-5\">5</a>]. In case of natural selection, selective removal of deleterious alleles occurs via negative selection or purifying selection [<a href=\"#r-6\">6</a>]. Purging by genetic drift can remove primarily deeply recessive alleles (may be deleterious or beneficial), whereas natural selection can remove any type of deleterious alleles [<a href=\"#r-7\">7</a>]. In small population, fixation of any selection pressures occurs promptly in fewer generations than that in large random mating populations. In strong purifying or negative selection on a locus, the purging of deleterious variants result in the incidental removal of linked variation. This occasional purging of non-deleterious alleles adjacent to deleterious ones is called background selection which may produce decreased level of variation surrounding the locus under selection [<a href=\"#r-8\">8</a>].<br />\r\nDue to degeneracy of genetic code, preference of codon usage determines the functional expression of a protein in an organism. The patterns of codon usage vary in different protein coding genes of a mitochondrial genome. Mutation pressure is dominant in determining codon usage patterns in mitochondrial genes that can be influenced by translational selection [<a href=\"#r-1\">1</a>]. Additional factors including compositional constraint of nucleotides, interaction between codon and anticodon, conservation of amino acids, etc., might drive the codon frequency, codon usage biasness in the genome of an organism [<a href=\"#r-9\">9</a>].<br />\r\nA plethora of manmade and natural causes have imposed a significant and alarming threat to the existence of various fish species in Bangladesh including <em>Cirrhinus reba</em>. The <em>C. reba</em> is a freshwater labeonine fish which is categorized as a near threatened (NT) species in Bangladesh [<a href=\"#r-10\">10-11</a>]. In the present study, base composition and skewness, various gene parameters, codon frequency and relative synonymous codon usage (RSCU), selection pressure and mutation rate have been analyzed for the protein coding genes of mitochondrial genome of the threatened labeonine fish, <em>C. reba</em> to provide valuable information on codon usage patterns and mutation pressure prevailing on the fish species.</p>"
},
{
"section_number": 2,
"section_title": "MATERIALS AND METHODS",
"body": "<p><strong>Base composition and skewness analysis</strong><br />\r\nBase composition of the complete mitochondrial genome of <em>C. reba</em> was analyzed by retrieving NCBI accession, MN862482 [<a href=\"#r-11\">11</a>]. The skewness [<a href=\"#r-12\">12</a>] of the base composition of any DNA sequences was measured by using the formula where the relative number of As is to Ts (AT skew = [A – T]/[A + T]) and the relative number of Gs is to Cs (GC skew = [G – C]/[G + C]). Skewness due to any particular nucleotides is attributed to differential mutational pressures originated from the asymmetric replication of mtDNA [<a href=\"#r-13\">13</a>]. CAIcal is a web server, freely available at http://genomes.urv.es/CAIcal, which was used to estimate important computational gene parameters. Some of the gene parameters are codon adaptation index (CAI) [<a href=\"#r-14\">14</a>], the effective number of codon (ENc) [<a href=\"#r-15\">15</a>] or G plus C percentage. Measurement of codon adaption index (CAI) is commonly applied to estimate codon usage bias in gene expression [<a href=\"#r-14\">14</a>]. CAI values range from 0 to 1, where higher values indicate a higher percentage of the most available codons.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Effective number of codon (ENc)</strong><br />\r\nThe ‘effective number of codon’ [<a href=\"#r-15\">15-16</a>] was estimated to identify the independent synonymous codon bias found in the genes of a genome. It indicates to what extent all 61 codons are engaged to express a gene. Expressivity of a gene depends on the efficient execution of codon usage [<a href=\"#r-17\">17</a>], for instance, most available codons and codons interacting to abundant tRNA species in the cells are preferred in case of highly expressed genes [<a href=\"#r-18\">18</a>]. Approaching the effective number of codons toward 20 imply that the respective gene is biased with its codons, while an unbiased gene approaches toward 61.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong> </strong><strong>Codon usage and RSCU analysis</strong><br />\r\nThe codon usage frequency was estimated by using MEGA7 [<a href=\"#r-19\">19</a>] and relative synonymous codon usage values (RSCU) were determined by using the formula described by Sharp et al. [<a href=\"#r-20\">20</a>]. The RSCU value of a codon is the ratio of the observed occurrences of a codon to that of the expected usage frequency under the assumption that all synonymous codons for a particular amino acid participate equally [<a href=\"#r-20\">20</a>; <a href=\"#r-21\">21</a>]. When all the synonymous codons of an amino acid participate without any preferences, the RSCU value of each codon equals to one. The codons with RSCU values >1.6, are considered to be “overrepresented” whereas the codons with RSCU values <0.6, are considered to be “underrepresented” [<a href=\"#r-21\">21; 22</a>].</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Selection pressure and mutation rate analysis</strong><br />\r\nFor exploring the mechanisms of DNA sequence evolution of protein-coding genes, a powerful computational tool is applied to estimate nonsynonymous (substitutions that replace an amino acid by another) and synonymous (silent) substitution rates in the coding sequences, termed as Ka and Ks, or dN and dS, respectively [<a href=\"#r-23\">23</a>]. <strong><em> </em></strong>Ka or dN reflects nonsynonymous substitutions per nonsynonymous site, and Ks or dS reflect synonymous substitutions per synonymous site. However, the Ka/Ks ratio (or ω, dN/dS) is frequently used as an estimator of selective strength for the evolution of DNA sequence, with ω > 1 implying adaptive (positive) selection, ω < 1 implying purifying (negative) or conservation selection, and ω close to 1 implying neutral mutation. In this study, dN/dS ratios were calculated by using an online based Synonymous Non-synonymous Analysis Program (SNAP v2.1.1) (www.hiv.lanl.gov) [<a href=\"#r-24\">24</a>] where sequences of the protein coding genes were uploaded after being aligned by ClustalW.</p>"
},
{
"section_number": 3,
"section_title": "RESULTS",
"body": "<p><strong>Base composition and AT-/GC-skewness</strong></p>\r\n\r\n<p>Based on the base composition of complete mitogenome of <em>C. reba</em>, the AT-/GC-skews analysis showed an overrepresentation of A and C resulting a lower number of T and G bases, respectively, where AT-skew was slightly positive (0.116) and GC-skew was slightly negative (-0.261) (<a href=\"#figure1\">Figure 1</a>). Except for ND6, all protein coding genes (PCGs) showed negative GC-skew, that indicated the higher occurrence of Cs. On the other hand, the COX1, ND3 and ND4L had slightly negative AT-skew (-0.03) and the ND6 had a negative AT-skewness of -0.528, meaning that the higher occurrence of Ts over As.</p>\r\n\r\n<div id=\"figure1\">\r\n<figure class=\"image\"><img alt=\"\" height=\"246\" src=\"/media/article_images/2023/53/26/178-1652598688-Figure1.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 1. </strong>Graphical presentation of AT- and GC skew of 13 PCGs of <em>Cirrhinus reba</em>.</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Gene parameters</strong><br />\r\nCodon adaption index (CAI) of the protein coding genes of the studied fish ranged from 0.56 to 0.641 implied that they experienced an average codon usage bias to be used in gene expression (<a href=\"#Table-1\">Table 1</a>). Effective number of codon (ENc) varied from 32.2 to 41.8, where ND1 (32.8), ATP8 (33.1) and CYTB (32.2) possessed the ENc values lower than 35 (<a href=\"#Table-1\">Table 1</a>). Percentage of GC bases at the 1<sup>st</sup>, 2<sup>nd</sup>, 3<sup>rd</sup> position of codons were shown in <a href=\"#figure2\">Figure 2</a>, where % GC3 was lower than % GC1 and % GC2 across all the PCGs.</p>\r\n\r\n<div id=\"Table-1\">\r\n<p><a href=\"https://jabet.bsmiab.org/table/178-1652598688-table1/\">Table-1</a><strong>Table 1. </strong>Gene parameters for 13 protein coding genes (PCGs) of the mitogenome of <em>C. reba.</em></p>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<div id=\"figure2\">\r\n<figure class=\"image\"><img alt=\"\" height=\"368\" src=\"/media/article_images/2023/53/26/178-1652598688-Figure2.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 2.</strong> Percentages of GC content at the 1st, 2nd and 3rd position of codons of 13 PCGs of <em>Cirrhinus reba</em>.</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Codon usage patterns</strong><br />\r\nA total of 3802 codons were used for coding 20 amino acids by a standard set of 64 codons. The amino acids Leucine and Serine were encoded by six different codons, whereas rest of the amino acids was encoded by either two or four codons. The hydrophobic amino acid Lue (16.44 %) was utilized at the highest frequency followed by hydrophobic amino acids, Ala (8.68 %) and Ile/Thr (8.02 %), whereas the frequency of the hydrophilic amino acid, Cys (0.66 %) was the lowest among the 13 PCGs (<a href=\"#figure3\">Figure 3A</a>). The most frequently used codon was CTA (7.26 %, Leu) followed by ATT (5.29 %, Ile) whereas, the rarely used codons were CGG (Arg), AGG (Lys), TCG (Ser), and TGG (Trp). The relative synonymous codon usage (RSCU) analysis revealed a total of 25 RSCU values with greater than 1.0 (Figure 3B). Several codons, GCA (Ala), CGA (Arg), CAA (Gln), GAA (Glu), GGA (Gly), CTA (Leu), TTA (Leu), AAA (Lys), CCA (Pro), TCA (Ser), ACA (Thr) and GTA (Val) were overpresented whereas the codons, GCG (Ala), CGG (Arg), GAT (Asp), CAG (Gln), CAC (His), TTG (Leu), CTG (Leu), AAG (Lys), ATG (Met), CCG (Pro), CCT (Pro), TCG (Ser), AGT (Ser), ACG (Thr), TGG (Trp), GTG (Val) and GTC (Val) were underpresented for coding the PCGs of the <em>C. reba</em> mitogenome.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Evolution rates of PCGs</strong><br />\r\nSequences of the protein coding genes of <em>C. reba</em> were retrieved and evolution rates were estimated referring to the mitogenomes of its phylogenetic relatives, <em>Labeo bata</em> (GenBank accession no.: AP011198.1) and <em>Labeo calbasu</em> (GenBank accession no.: AP012143.1). The estimated Ka/Ks ratios also known as dN/dS ratios between <em>C. reba </em>and <em>L. bata </em>ranged from the lowest value (0.008) for COX1 gene to the highest value (0.997) for ND1 gene whereas, the values between <em>C. reba </em>and <em>L. calbasu </em>ranged from the lowest value (0.008) for COX3 gene to the highest value (0.673) for ND1 gene (<a href=\"#figure4\">Figure 4</a>). The ND1 gene showed the higher Ka/Ks ratios comparing to that of other protein coding genes among the three fish species.</p>\r\n\r\n<div id=\"figure3\">\r\n<figure class=\"image\"><img alt=\"\" height=\"704\" src=\"/media/article_images/2023/53/26/178-1652598688-Figure3.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 3.</strong> Codon usage frequency (A) and relative synonymous codon usage (RSCU) (B) of the mitochondrial genome of <em>Cirrhinus reba</em> (Reverse complementary sequence was used for ND6 gene which was encoded on the L-strand and incomplete stop codons were not included).</figcaption>\r\n</figure>\r\n</div>\r\n\r\n<div id=\"figure4\">\r\n<figure class=\"image\"><img alt=\"\" height=\"291\" src=\"/media/article_images/2023/53/26/178-1652598688-Figure4.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 4. </strong>Selective strength of protein coding genes of <em>Cirrhinus reba</em> (CR) mitogenome comparing to that of <em>Labeo bata</em> (LB) (GenBank accession no.: AP011198.1) and <em>Labeo calbasu</em> (LC) (GenBank accession no: AP012143.1). The ratio indicates the rate of non-synonymous substitutions to the rate of synonymous substitutions (Ka/Ks) for each protein coding genes.</figcaption>\r\n</figure>\r\n\r\n<p> </p>\r\n</div>"
},
{
"section_number": 4,
"section_title": "DISCUSSION",
"body": "<p>Nucleotide skew may be attributed to the equilibrium between mutation and selection pressures which are known to be affected by asymmetric mechanisms, largely of mtDNA replication, repair and to a lesser extent of transcription [<a href=\"#r-25\">25,26</a>] which inferring the functionality of related genes. AT-skewness and GC-skewness are considered as a vital indicator to analyze strand asymmetry and the patterns of nucleotide composition of DNA sequences [<a href=\"#r-27\">27</a>]. The lowest AT-skewness and highest GC-skewness value were observed in case of ND6, which was the only PCG exhibiting a positive value in the GC-skew curve and was common to other reports [<a href=\"#r-26\">26, 28, 29</a>]. Mitochondrial genomes exhibit a strong strand-specific compositional bias [<a href=\"#r-30\">30</a>] where heavy strand (H-strand) is guanine rich and light-strand (L- strand) is guanine poor [<a href=\"#r-31\">31</a>]. In our analysis, ND6 which was located on the L- strand, showed larger fluctuation in AT/GC-skew value, suggesting that the selection and differential mutational pressure on this gene might be significantly different from other genes as reported in other fish species [<a href=\"#r-26\">26</a>].<br />\r\nThe second codon positions were reported with the over-representation of Ts in [<a href=\"#r-11\">11</a>] and the coded amino acids might be related to a preference for nonpolar and hydrophobic amino acids in membrane-associated proteins [<a href=\"#r-30\">30</a>]. The codons which possessed a T at the second position are coded for valine, liucine, isoliucine, alanine, phenylalanine, and methionine. These are nonpolar and hydrophobic amino acids [<a href=\"#r-32\">32; 33</a>]. Codons ending with G or C at the 3<sup>rd</sup> position might have compositional constraint under mutation pressure. However, PCGs of <em>C. reba </em>might compromise with this type of constraint enabling higher mutation rates.<br />\r\nCodon usage pattern is exploited for those codons which are more quickly and efficiently translated into protein by the cellular machineries of an organism. However, translational selection is demonstrated in case of highly expressed genes by engaging codon usage patterns. The preferred codons usually occur at higher frequencies in the sequence of highly expressed genes than that in weakly expressed genes [<a href=\"#r-1\">1</a>]. Recently, 30 RSCU values were reported in a cottoid fish [<a href=\"#r-13\">13</a>]. In the present study, a total of 25 RSCU values were identified with the values of greater than 1. Besides, 12 codons were revealed as “overpresented” that implied to engage in highly expressed genes for efficient protein synthesis via translational selection.<br />\r\nAll the 13 PCGs of three croakers were investigated for mutation rates in a study, where the Ka/Ks ratios ranged from 0.0100 to 0.2714 [<a href=\"#r-13\">13</a>]. In that study, the Ka/Ks ratios indicated that the protein coding genes of those three fish species evolved under strong purifying selection [<a href=\"#r-13\">13</a>]. Evolution of a DNA sequence is driven by non-adaptive forces including genetic drift and mutation pressure [<a href=\"#r-34\">34</a>]. Under purifying selection, although, it is difficult to establish any mutations driven demerits, the selection pressures posed differently among the genes and are supposed to evolve in different ways [<a href=\"#r-34\">34</a>]. Ka/Ks ratio between <em>C. reba </em>and <em>L. bata </em>was close to one which meant neutral mutation in ND1 gene of <em>C. reba</em> implying neither beneficial nor detrimental to its survival or reproduction. Except for ND1, COX2 and ND5, all the values of Ka/Ks ratios were less than 0.1. However, we can consider that the mutation occurred in these PCGs of <em>C. reba </em>was driven by purifying or negative selection. Negative selection indicates prevalence of more synonymous mutations than non-synonymous mutations where ancestral state and/or more diverse gene pool is essential to conserve for appropriate functions of encoded proteins. If negative selection is unable to remove the harmful mutations, an accumulation of deleterious mutations occurs [<a href=\"#r-6\">6</a>]. If the negative selection is too strong in the whole population or hard selection occurs that reduce the number of surviving offspring, the possibility of extinction will increase. Climate changes and/or habitat alterations are responsible for threatening of those species under strong negative selection.</p>"
},
{
"section_number": 5,
"section_title": "CONCLUSION",
"body": "<p>The mutation in PCGs of <em>C. reba</em> dominated the synonymous substitutions among its relatives which might contribute to the genetic stability and diversity of this threatened labeonine fish. The overpresenting codons indicating the functions of genes under translational selection might influence the life cycle of the fish species. Research works should be conducted linking the environmental conditions and nutritional requirements along with the major causes of threatened/endangered state of the fish species. For the conservation of the populations of <em>C. reba</em>, different strategies should be considered, e.g., <em>in situ </em>conservation by installing fish sanctuary, <em>ex situ</em> conservation by adopting artificial breeding program, stopping, or prohibiting indiscriminate use of insecticides, fertilizers in agricultural fields, practices of bioremediation, treating industrial effluents before mixing to the water bodies for decreasing pollution of their habitats and breeding grounds.</p>"
},
{
"section_number": 6,
"section_title": "ACKNOWLEDGEMENTS",
"body": "<p>We are highly thankful to the assigned reviewers for their constructive critiques, suggestions, and comments to improve the manuscript.</p>"
},
{
"section_number": 7,
"section_title": "AUTHOR CONTRIBUTIONS",
"body": "<p>MNI designed the outlines of the manuscript and drafted the original manuscript. MNI and SS performed the formal analysis of the data. MNI and SS wrote, revised and edited the manuscript.</p>"
},
{
"section_number": 8,
"section_title": "CONFLICTS OF INTEREST",
"body": "<p>There is no conflict of interest among the authors.</p>"
}
],
"figures": [
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/53/26/178-1652598688-Figure1.jpg",
"caption": "Figure 1. Graphical presentation of AT- and GC skew of 13 PCGs of Cirrhinus reba.",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/53/26/178-1652598688-Figure2.jpg",
"caption": "Figure 2. Percentages of GC content at the 1st, 2nd and 3rd position of codons of 13 PCGs of Cirrhinus reba.",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/53/26/178-1652598688-Figure3.jpg",
"caption": "Figure 3. Codon usage frequency (A) and relative synonymous codon usage (RSCU) (B) of the mitochondrial genome of Cirrhinus reba (Reverse complementary sequence was used for ND6 gene which was encoded on the L-strand and incomplete stop codons were not included).",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/53/26/178-1652598688-Figure4.jpg",
"caption": "Figure 4. Selective strength of protein coding genes of Cirrhinus reba (CR) mitogenome comparing to that of Labeo bata (LB) (GenBank accession no.: AP011198.1) and Labeo calbasu (LC) (GenBank accession no: AP012143.1). The ratio indicates the rate of non-synonymous substitutions to the rate of synonymous substitutions (Ka/Ks) for each protein coding genes.",
"featured": false
}
],
"authors": [
{
"id": 478,
"affiliation": [
{
"affiliation": "Department of Biotechnology, Sher-e-Bangla Agricultural University, Sher-e-Bangla Nagar, Dhaka-1207, Bangladesh"
}
],
"first_name": "Mohammad Nazrul",
"family_name": "Islam",
"email": "nazrul.sau.bd@gmail.com",
"author_order": 1,
"ORCID": "http://orcid.org/0000-0001-5692-3566",
"corresponding": true,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "Mohammad Nazrul Islam, PhD; Department of Biotechnology, Shere Bangla Agricultural University, Sher-e-Bangla Nagar, Bangladesh, e-mail: nazrul.sau.bd@gmail.com",
"article": 119
},
{
"id": 479,
"affiliation": [
{
"affiliation": "Fisheries Biotechnology Division, National Institute of Biotechnology, Dhaka-1349, Bangladesh"
}
],
"first_name": "Shirin",
"family_name": "Sultana",
"email": null,
"author_order": 2,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 119
}
],
"views": 1417,
"downloads": 243,
"references": [
{
"id": 3846,
"serial_number": 1,
"pmc": null,
"reference": "Jia W, Higgs PG. Codon usage in mitochondrial genomes: distinguishing context-dependent mutation from translational selection. Mol Biol Evol 2008; 25(2):339-51.",
"DOI": null,
"article": 119
},
{
"id": 3847,
"serial_number": 2,
"pmc": null,
"reference": "Urbina D, Tang B, Higgs PG. The response of amino acid frequencies to directional mutation pressure in mitochondrial genome sequences is related to the physical properties of the amino acids and to the structure of the genetic code. J Mol Evol 2006; 62(3):340-61.",
"DOI": null,
"article": 119
},
{
"id": 3848,
"serial_number": 3,
"pmc": null,
"reference": "Faith JJ, Pollock DD. Likelihood analysis of asymmetrical mutation bias gradients in vertebrate mitochondrial genomes. Genet 2003; 165(2):735-45.",
"DOI": null,
"article": 119
},
{
"id": 3849,
"serial_number": 4,
"pmc": null,
"reference": "Billington N, Hebert PD. Mitochondrial DNA diversity in fishes and its implications for introductions. Canadian J Fish Aquat Sci 1991; 48(S1):80-94.",
"DOI": null,
"article": 119
},
{
"id": 3850,
"serial_number": 5,
"pmc": null,
"reference": "Kimura M. Evolutionary rate at the molecular level. Nature publishing group [Internet], 1968; 217 (5129): 624–626.",
"DOI": null,
"article": 119
},
{
"id": 3851,
"serial_number": 6,
"pmc": null,
"reference": "Loewe L. Negative Selection. Nature Education 2008; 1 (1): 59.",
"DOI": null,
"article": 119
},
{
"id": 3852,
"serial_number": 7,
"pmc": null,
"reference": "Glémin S. How are deleterious mutations purged? Drift versus nonrandom mating. Evolution 2003; 57(12):2678-87.",
"DOI": null,
"article": 119
},
{
"id": 3853,
"serial_number": 8,
"pmc": null,
"reference": "Charlesworth B, Morgan MT, Charlesworth D. The effect of deleterious mutations on neutral molecular variation. Genet 1993; 134(4):1289-303.",
"DOI": null,
"article": 119
},
{
"id": 3854,
"serial_number": 9,
"pmc": null,
"reference": "Sahoo S, Das SS, Rakshit R. Codon usage pattern and predicted gene expression in Arabidopsis thaliana. Gene 2019; 721:100012.",
"DOI": null,
"article": 119
},
{
"id": 3855,
"serial_number": 10,
"pmc": null,
"reference": "Bangladesh IUCN. Red List of Bangladesh Volume 5: Freshwater Fishes. IUCN, International Union for Conservation of Nature, Bangladesh Country Office, Dhaka, Bangladesh. 2015; 16:360.",
"DOI": null,
"article": 119
},
{
"id": 3856,
"serial_number": 11,
"pmc": null,
"reference": "Islam MN, Sultana S, Alam MJ. Sequencing and annotation of the complete mitochondrial genome of a threatened labeonine fish, Cirrhinus reba. Genomics Inform 2020; 18(3). e32. doi: 10.5808/GI.2020.18.3.e32.",
"DOI": null,
"article": 119
},
{
"id": 3857,
"serial_number": 12,
"pmc": null,
"reference": "Perna NT, Kocher TD. Patterns of nucleotide composition at fourfold degenerate sites of animal mitochondrial genomes. J Mol Evol 1995; 41(3):353-8.",
"DOI": null,
"article": 119
},
{
"id": 3858,
"serial_number": 13,
"pmc": null,
"reference": "Yang H, Xia J, Zhang JE, Yang J, Zhao H, Wang Q, Sun J, Xue H, Wu Y, Chen J, Huang J. Characterization of the complete mitochondrial genome sequences of three croakers (perciformes, sciaenidae) and novel insights into the phylogenetics. Int J Mol Sci 2018; 19(6):1741.",
"DOI": null,
"article": 119
},
{
"id": 3859,
"serial_number": 14,
"pmc": null,
"reference": "Sharp PM, Li WH. The codon adaptation index-a measure of directional synonymous codon usage bias, and its potential applications. Nucleic Acids Res 1987; 15(3):1281-95.",
"DOI": null,
"article": 119
},
{
"id": 3860,
"serial_number": 15,
"pmc": null,
"reference": "Wright F. The ‘effective number of codons’ used in a gene. Gene 1990; 87(1):23-9.",
"DOI": null,
"article": 119
},
{
"id": 3861,
"serial_number": 16,
"pmc": null,
"reference": "Fuglsang A. The ‘effective number of codons’ revisited. Biochem Biophys Res Commun 2004; 317(3):957-64.",
"DOI": null,
"article": 119
},
{
"id": 3862,
"serial_number": 17,
"pmc": null,
"reference": "Gouy M, Gautier C. Codon usage in bacteria: correlation with gene expressivity. Nucleic Acids Res 1982; 0(22):7055-74.",
"DOI": null,
"article": 119
},
{
"id": 3863,
"serial_number": 18,
"pmc": null,
"reference": "Ikemura T. Codon usage and tRNA content in unicellular and multicellular organisms. Mol Biol Evol 1985; 2(1):13-34.",
"DOI": null,
"article": 119
},
{
"id": 3864,
"serial_number": 19,
"pmc": null,
"reference": "Kumar S, Stecher G, Tamura K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol Biol Evol 2016; 33:1870–1874.",
"DOI": null,
"article": 119
},
{
"id": 3865,
"serial_number": 20,
"pmc": null,
"reference": "Sharp PM, Tuohy TM, Mosurski KR. Codon usage in yeast: cluster analysis clearly differentiates highly and lowly expressed genes. Nucleic Acids Res 1986; 14(13):5125-43.",
"DOI": null,
"article": 119
},
{
"id": 3866,
"serial_number": 21,
"pmc": null,
"reference": "He Z, Gan H, Liang X. Analysis of synonymous codon usage bias in potato virus M and Its adaption to hosts. Viruses 2019; 11(8):752.",
"DOI": null,
"article": 119
},
{
"id": 3867,
"serial_number": 22,
"pmc": null,
"reference": "Gupta SK, Ghosh TC. Gene expressivity is the main factor in dictating the codon usage variation among the genes in Pseudomonas aeruginosa. Gene 2001; 273(1):63-70.",
"DOI": null,
"article": 119
},
{
"id": 3868,
"serial_number": 23,
"pmc": null,
"reference": "Hurst LD. The Ka/Ks ratio: diagnosing the form of sequence evolution. Trends Genet: TIG 2002; 18(9):486-.",
"DOI": null,
"article": 119
},
{
"id": 3869,
"serial_number": 24,
"pmc": null,
"reference": "Korber B. HIV Signature and Sequence Variation Analysis. In: Rodrigo AG and Learn GH (eds). Computational Analysis of HIV Molecular Sequences. Kluwer Academic Publishers: Dordrecht, Netherlands, 2000, pp 55-72.",
"DOI": null,
"article": 119
},
{
"id": 3870,
"serial_number": 25,
"pmc": null,
"reference": "Fonseca MM, Harris DJ, Posada D. The inversion of the control region in three mitogenomes provides further evidence for an asymmetric model of vertebrate mtDNA replication. PLoS One 2014; 9(9):e106654.",
"DOI": null,
"article": 119
},
{
"id": 3871,
"serial_number": 26,
"pmc": null,
"reference": "Ruan H, Li M, Li Z, Huang J, Chen W, Sun J, Liu L, Zou K. Comparative analysis of complete mitochondrial genomes of three Gerres fishes (Perciformes: Gerreidae) and primary exploration of their evolution history. Int J Mol Sci 2020; 21(5):1874.",
"DOI": null,
"article": 119
},
{
"id": 3872,
"serial_number": 27,
"pmc": null,
"reference": "Lü Z, Zhu K, Jiang H, Lu X, Liu B, Ye Y, Jiang L, Liu L, Gong L. Complete mitochondrial genome of Ophichthus brevicaudatus reveals novel gene order and phylogenetic relationships of Anguilliformes. Int J Biol Macromol 2019; 135:609-18.",
"DOI": null,
"article": 119
},
{
"id": 3873,
"serial_number": 28,
"pmc": null,
"reference": "Mu X, Liu Y, Lai M, Song H, Wang X, Hu Y, Luo J. Characterization of the Macropodus opercularis complete mitochondrial genome and family Channidae taxonomy using Illumina-based de novo transcriptome sequencing. Gene 2015; 559(2):189-95.",
"DOI": null,
"article": 119
},
{
"id": 3874,
"serial_number": 29,
"pmc": null,
"reference": "Boore JL. Requirements and standards for organelle genome databases. Omics: J Integ Biol 2006; 10(2):119-26.",
"DOI": null,
"article": 119
},
{
"id": 3875,
"serial_number": 30,
"pmc": null,
"reference": "Asakawa S, Kumazawa Y, Araki T, Himeno H, Miura KI, Watanabe K. Strand-specific nucleotide composition bias in echinoderm and vertebrate mitochondrial genomes. J Mol Evol 1991; 32(6):511-20.",
"DOI": null,
"article": 119
},
{
"id": 3876,
"serial_number": 31,
"pmc": null,
"reference": "Anderson S, Bankier AT, Barrell BG, de Bruijn MH, Coulson AR, Drouin J, Eperon IC, Nierlich DP, Roe BA, Sanger F, Schreier PH. Sequence and organization of the human mitochondrial genome. Nature 1981; 290(5806):457-65.",
"DOI": null,
"article": 119
},
{
"id": 3877,
"serial_number": 32,
"pmc": null,
"reference": "Junqueira AC, Lessinger AC, Torres TT, da Silva FR, Vettore AL, Arruda P, Espin AM. The mitochondrial genome of the blowfly Chrysomya chloropyga (Diptera: Calliphoridae). Gene 2004; 339:7-15.",
"DOI": null,
"article": 119
},
{
"id": 3878,
"serial_number": 33,
"pmc": null,
"reference": "Helfenbein KG, Brown WM, Boore JL. The complete mitochondrial genome of the articulate brachiopod Terebratalia transversa. Mol Biol Evol 2001; 18(9):1734-44.",
"DOI": null,
"article": 119
},
{
"id": 3879,
"serial_number": 34,
"pmc": null,
"reference": "Lynch M, Koskella B, Schaack S. Mutation pressure and the evolution of organelle genomic architecture. Science 2006; 311(5768):1727-30.",
"DOI": null,
"article": 119
}
]
},
{
"id": 116,
"slug": "178-1651419347-granzyme-b-gene-polymorphisms-and-risk-of-hepatocellular-carcinoma-in-patients-with-chronic-hepatitis",
"featured": false,
"slider": false,
"issue": "Vol5 Issue3",
"type": "original_article",
"manuscript_id": "178-1651419347",
"recieved": "2022-05-12",
"revised": null,
"accepted": "2022-06-20",
"published": "2022-07-06",
"pdf_file": "https://jabet.bsmiab.org/media/pdf_file/2023/07/178-1651419347.pdf",
"title": "Granzyme B gene polymorphisms and risk of hepatocellular carcinoma in patients with chronic hepatitis",
"abstract": "<p>Infection with the hepatitis B virus (HBV) continues to be a hazard for public health across the globe. Chronic hepatitis, cirrhosis, and hepatocellular carcinoma (HCC) are all possible outcomes. It is obvious that certain patients with chronic hepatitis B (CHB) viral infection developed HCC, while other under almost similar circumstances do not. The present study aimed to investigate the possible a link between three single nucleotide gene polymorphisms (SNPs) in GzmB genes with the development of HCC. A total of 85 patients diagnosed with CHB participated in this research (40 patients with HCC and 45 patients without HCC). Three SNPs in GzmB gene (rs7144366, rs8192917 and rs2236338) were genotyped using restriction fragment length polymorphism (RFLP). The haplotype blocks derived from the three SNPs were assembled, and the linkage disequilibrium (LD) between the SNPs was determined using the SHEsis software. The homozygous mutant genotype (CC) was shown to be significantly more common in patients with HCC (27.5 %) than in those without HCC (11.11 %) (OR= 3.93, 95% CI=1.13-13.62, p=0.031). At allelic level, the mutant allele (C) was more frequent in patients with HCC than those without HCC (46.25% vs. 26.67%) with a significant deviation (OR=2.36, 95%CI= 1.25- 4.49, p= 0.008). The haplotype block CCG was more common among patients with HCC (26.25%) than those without HCC (12.22%) with a significant difference (OR= 2.56, 95%= 1.14-5.71, p= 0.022). The study indicated that individuals carrying the mutant homozygous (CC) of the SNP rs8192917 and allele C of this SNP may have a higher chance of developing HCC compared with those carrying other genotypes and T allele of the SNP. The haplotype block CCG (corresponding for C allele of rs7144366, C allele of rs8192917 and G allele of rs2236338) might be regarded as a risk factor for the emergence of HCC in patients with CHB.</p>",
"journal_reference": "J Adv Biotechnol Exp Ther. 2022; 5(3): 577-589.",
"academic_editor": "Md Jamal Uddin, PhD; ABEx Bio-Research Center, Dhaka-1230, Bangladesh",
"cite_info": "Alshamary HS, Mayah QSA, et al. Granzyme B gene polymorphisms and risk of hepatocellular carcinoma in patients with chronic hepatitis. J Adv Biotechnol Exp Ther. 2022; 5(3): 577-589.",
"keywords": [
"Chronic hepatitis B",
"Polymorphism",
"Hepatocellular carcinoma",
"Granzyme B"
],
"DOI": "doi.org/10.5455/jabet.2022.d137",
"sections": [
{
"section_number": 1,
"section_title": "INTRODUCTION",
"body": "<p>Approximately 90% of all initial liver malignancies are triggered by hepatocellular carcinoma (HCC), prevalent in the world’s most populated areas. It accounts for 9.2 % of all new cancer cases and the third biggest cause of cancer-related death, and it is the fifth most prevalent cancer in men and the seventh most common cancer in women in the world, following only stomach and lung cancers [<a href=\"#r-1\">1</a>]. The activation of various cellular receptors (such as toll-like receptors (TLR)-3, TLR-9, retinoic acid-inducible gene (RIG)-1) results in the rapid production of antiviral cytokines such as interferon (IFN), which, in cooperation with natural killer (NK) cells, limits the initial spread of the hepatitis B virus (HBV) [<a href=\"#r-2\">2</a>]. The humeral arm of immune response may also participate in body defense against the virus through the maturation and proliferation of B-cells to create antibodies specific to hepatitis B antigen [<a href=\"#r-3\">3</a>]. The granules of cytotoxic T lymphocyte (CTL) and NK cells are discharged into the immunological synapse that has been established with their target cells. Serine protease granzymes (Gzms) are the granules’ most potent contents, which are transported into the cytoplasm of the target cells by the pore-forming protein perforin and trigger programmed cell death [<a href=\"#r-4\">4</a>]. Granzyme B is the most important member of granzymes. It causes early DNA damage by causing the death of target cells through a mitochondria-dependent mechanism [<a href=\"#r-5\">5</a>]. CTL responds to viruses infected cells by producing Gzms. Those Gzms should work at maximum speed with lower Km and should be free of the inhibitors for the enzymes to initiate apoptosis successfully [<a href=\"#r-3\">3</a>]. The objective of the current investigation was to investigate the possibility of a relationship between three single nucleotide gene polymorphisms in the GzmB gene and the development of HCC in patients with CHB.</p>"
},
{
"section_number": 2,
"section_title": "MATERIALS AND METHODS",
"body": "<p><strong>Study population</strong><br />\r\nThis study was conducted from 1 January to 31 December 2021, in the Department of Chemistry and Biochemistry at Al-Nahrain University’s College of Medicine, in collaboration with the Department of Biochemistry Laboratory at Al-Imamain Alkadhimain Teaching Hospital and Medical Research Unit/College of Medicine/Al-Nahrain University. The Institutional Review Board (IRB) of the College of Medicine at Al-Nahrain University gave its clearance to the research project (number 202012158 on 19 January 2021). A total of 85 patients with CHB were included in the study. Those patients were split up into two different groups. Group 1 included 45 patients diagnosed with hepatocellular carcinoma, and group 2 included 45 patients without hepatocellular carcinoma.<br />\r\nThe diagnosis of CHB was primarily based on positivity for HBsAg and anti-HBc-IgG antibodies accompanied by viral load beyond the reference range. The diagnosis of HCC depended on ultrasound (US) findings and Computed tomography (CT) scan. Patients with renal failure, connective tissue disease, acute and chronic infection, hepatitis C virus (HCV), and other malignancies were excluded from the study.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Blood samples</strong><br />\r\nVenous blood (about 5 ml) was collected from all subjects, with 2 mL in an EDTA tube and 3 mL in a plan tube. The serum was extracted from the latter after centrifugation and stored at -20 ºC until used.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>HBV serological markers</strong><br />\r\nReady commercial kits were used to detect HBsAg, anti-HBc IgG, HBeAg, and anti-HBe using the commercial ELISA technique (Bioelisa/biokit/Barcelona/Spain).</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>DNA extraction and gene amplification</strong><br />\r\nFollowing the manufacturer’s instructions, a ready kit (gSYNCTM DNA Mini Kit Whole Blood Protocol/ Geneaid/ Korea) was used to extract DNA from blood samples. The GzmB gene fragments involving rs7144366, rs8192917, and rs2236338 gene polymorphisms were amplified using three sets of primers (<a href=\"#Table-1\">Table 1</a>).<br />\r\nThe final concentration of 100 pmole/L was achieved by dissolving the primers in DNase and RNase free water, which was used in the experiments. Mixing 10 μL of the stock solution with 90 μL of distilled water resulted in a working solution with a concentration of 10 pmol/L. Each master mix tube included 4 μL of template DNA and 2 μL of primers. The amplification of GzmB gene segments matching to the three SNPs was carried out under the circumstances of a flowing PCR reaction. The annealing temperature was the sole variable; it was 58, 60, and 61 <sup>o</sup>C for the rs7144366, rs8192917, and rs2236338 gene polymorphisms, respectively. The first denaturation was performed at 95<sup>o</sup>C for 5 minutes, followed by denaturation at 94<sup>o</sup>C for 30 seconds, annealing for 30 seconds, elongation at 72<sup> o</sup>C for 1 minute, and final elongation at 72 <sup>o</sup>C for 7 minutes.</p>\r\n\r\n<div id=\"Table-1\">\r\n<p><a href=\"https://jabet.bsmiab.org/table/178-1651419347-table1/\">Table-1</a><strong>Table 1. </strong>Primers and their corresponding single nucleotide polymorphisms.</p>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Restriction fragment length polymorphism</strong><br />\r\nThe PCR product was digested with the restriction enzyme. The restriction enzymes and their cutting sequence are illustrated in <a href=\"#Table-2\">Table 2</a>.<br />\r\nFive μL of PCR products were mixed with 0.1μL of the selected restriction enzyme (Sibenzyme, Russia). Restriction buffer (10X) 1.5 µL and bovine serum albumin (BSA) 0.1µL were added to the mixture. The reaction mixture was then completed to 15µ l by molecular grad water. Mineral oil (20 µl) was added to each tube to prevent evaporation. The reaction mixture was incubated in 60<sup>◦</sup> C water bath for 3 hours. Restriction products were resolved on 2% agarose.</p>\r\n\r\n<div id=\"Table-2\">\r\n<p><a href=\"https://jabet.bsmiab.org/table/178-1651419347-table2/\">Table-2</a><strong>Table 2.</strong> The restriction enzymes, their cutting sequence, and fragment length resulted from digestion.</p>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>PCR product sequencing</strong><br />\r\nTen percent of the PCR products were sent to the Macrogen Company in South Korea to verify RFLP for direct sequencing. The generated nucleotide sequences were put through a BLAST alignment process, which compared them to the reference sequence. This tool is accessible online at <a href=\"http://www.ncbi.nlm.nih.gov/\">http://www.ncbi.nlm.nih.gov</a>.</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong>Statistical analysis</strong><br />\r\nStatistical analysis was performed using SPSS 25.0 (SPSS, Chicago). The student t-test was used. The genotype divergence from Hardy-Weinberg Equilibrium (HWE) was assessed using Chi-square. The OR and 95% confidence interval (CI) were calculated using binary logistic regression (CI). SHEsis software was used to create haplotypes and calculate LD between the GzmB gene polymorphisms rs7144366, rs8192917, and rs2236338. A statistically significant difference was defined as a p-value less than 0.05.</p>"
},
{
"section_number": 3,
"section_title": "RESULTS",
"body": "<p><strong>Demographic and laboratory data of the study population</strong><br />\r\nAlthough patients with HCC demonstrated higher mean age than those without HCC (42.7±14.18 years vs. 37.8±13.46 years), the difference was not significant. Both groups were similar in their characteristics in terms of gender distribution and HBeAg positivity, with no significant differences. However, patients without HCC had far more frequent positivity for hepatitis B antigen and antibody (HBeAb) than those with HCC (46.67% vs. 7.5%), with a highly significant difference (<a href=\"#Table-3\">Table 3-5</a>). Interestingly, all patients in both groups were positive for HBsAg and HBcAb-IgG.</p>\r\n\r\n<div id=\"Table-3\">\r\n<p><a href=\"https://jabet.bsmiab.org/table/178-1651419347-table3/\">Table-3</a><strong>Table 3. </strong>Characteristics of the participants, both demographic and laboratory-based.</p>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Molecular assays</strong><br />\r\nThree SNPs were investigated in the present study for their association with HCC. The genotyping was performed by RFLP. The different genotypes in all included SNPs were in good accordance with HWE in both groups (with and without HCC).</p>\r\n\r\n<p> </p>\r\n\r\n<p><strong><em>rs7144366</em></strong><br />\r\nGel electrophoresis of the PCR product of rs7144366 polymorphism is shown in <a href=\"#figure1\">Figure 1</a>. The fragment length was 97bp. Digestion with SfcI restriction enzyme revealed three genotypes: TT, CT, and CC as shown in <a href=\"#figure2\">Figure 2</a> and <a href=\"#figure3\">3</a>.<br />\r\nAlthough the homozygous mutant genotype (CC) was more frequent in patients with HCC than those without HCC (27.5% vs. 13.33%), the difference was not significant. Likewise, a higher percentage of HCC patients had the allele C (45%) than those without HCC (35.56%), with no significant difference (<a href=\"#Table-4\">Table 4</a>).</p>\r\n\r\n<div id=\"figure1\">\r\n<figure class=\"image\"><img alt=\"\" height=\"147\" src=\"/media/article_images/2023/58/26/178-1651419347-Figure1.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 1. </strong>Gel electrophoresis (1% agarose) of rs7144366 gene polymorphism amplified with specific pair of primers using conventional PCR run for 45 min at 129 V. The PCR product was stained with ethidium bromide. The fragment length was 97bp.</figcaption>\r\n</figure>\r\n</div>\r\n\r\n<div id=\"figure2\">\r\n<figure class=\"image\"><img alt=\"\" height=\"170\" src=\"/media/article_images/2023/58/26/178-1651419347-Figure2.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 2.</strong> Genotyping of rs7144366 gene polymorphism after digestion with SfcI and stained with ethidium bromide. Lanes 1,3,6,7,8, and 11: TT; lanes 2,5, and 10: CT, lanes 4 and 9: CC genotype; M: DNA 50 bp DNA marker.</figcaption>\r\n</figure>\r\n</div>\r\n\r\n<div id=\"figure3\">\r\n<figure class=\"image\"><img alt=\"\" height=\"164\" src=\"/media/article_images/2023/58/26/178-1651419347-Figure3.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 3. </strong>Sequence analysis of the rs7144366, the reverse strand. The R in the upper frame represents the heterozygous genotype (TC), the G in the middle frame represents the mutant genotype (CC), and the A in the bottom frame represents the homozygous wild type of genotype (TT).</figcaption>\r\n</figure>\r\n</div>\r\n\r\n<div id=\"Table-4\">\r\n<p><a href=\"https://jabet.bsmiab.org/table/178-1651419347-table4/\">Table-4</a><strong>Table 4. </strong>The frequency of different genotypes and allele of rs7144366 in hepatitis patients with and without HCC.</p>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong><em>rs8192917</em></strong><br />\r\nGel electrophoresis of the PCR product of rs8192917 polymorphism is shown in <a href=\"#figure4\">Figure 4</a>. The fragment length was 420bp. Digestion with BsmA I restriction enzyme revealed three genotypes: TT, CT, and CC, as shown in <a href=\"#figure5\">Figure 5</a> and <a href=\"#figure6\">6</a>.<br />\r\nThe homozygous mutant genotype (CC) was far more frequent in patients with HCC (27.5%) than those without HCC (11.11%) with a significant difference (OR= 3.93, 95%CI=1.13-13.62, p=0.031). This polymorphism seems to have recessive inheritance as the CC+CT genotypes were more common in patients with HCC than those without HCC (65% vs. 42.22%) with a significant difference (OR= 2.54, 95%CI= 1.06-6.12). At allelic level, the mutant allele (C) was more frequent in patients with than those without HCC (46.25% vs. 26.67%) with a notable distinction (OR=2.36, 95%CI= 1.25- 4.49, p= 0.008) as shown in <a href=\"#Table-5\">Table 5</a>.</p>\r\n\r\n<div id=\"figure4\">\r\n<figure class=\"image\"><img alt=\"\" height=\"163\" src=\"/media/article_images/2023/58/26/178-1651419347-Figure4.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 4. </strong>Gel electrophoresis (1% agarose) of rs<em> 8192917</em> gene polymorphism amplified with specific pair of primers using conventional PCR run for 45 min at 129 V. The PCR product was stained with ethidium bromide. The fragment length was 420 bp.</figcaption>\r\n</figure>\r\n</div>\r\n\r\n<div id=\"figure5\">\r\n<figure class=\"image\"><img alt=\"\" height=\"161\" src=\"/media/article_images/2023/58/26/178-1651419347-Figure5.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 5. </strong>Genotyping of rs7144366 gene polymorphism after digestion with BsmA I and stained with ethidium bromide. Lanes 1,2,4,7,8,10,11, 13 and 14: TT; lanes 5,6 and 12: CT, lanes 4 and 9: CC genotype; M: 100 bp DNA maker.</figcaption>\r\n</figure>\r\n</div>\r\n\r\n<div id=\"figure6\">\r\n<figure class=\"image\"><img alt=\"\" height=\"181\" src=\"/media/article_images/2023/58/26/178-1651419347-Figure6.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 6. </strong>Sequence analysis of the rs8192917, the reverse strand. The homozygous wild type is represented by the letter A in the top frame (TT), the G in the second line of the frame represents the mutant genotype (CC), while the R in the lower line represents the heterozygous genotype (TC).</figcaption>\r\n</figure>\r\n</div>\r\n\r\n<div id=\"Table-5\">\r\n<p><a href=\"https://jabet.bsmiab.org/table/178-1651419347-table5/\">Table-5</a><strong>Table 5.</strong> The prevalence of various genotypes and alleles of rs8192917 in hepatitis patients with and without HCC.</p>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong><em>rs2236338</em></strong><br />\r\nGel electrophoresis of the PCR product of rs2236338 polymorphism is shown in <a href=\"#figure7\">Figure 7</a>. The fragment length was 400bp. Digestion with BsmF I restriction enzyme revealed three genotypes: AA, AG and GG as shown in <a href=\"#figure8\">Figure 8</a> and<a href=\"#figure9\"> 9</a>. There were no significant differences in the frequency of various genotypes and alleles of this polymorphism between individuals who had HCC and those who did not have the condition (<a href=\"#Table-6\">Table 6</a>).</p>\r\n\r\n<div id=\"figure7\">\r\n<figure class=\"image\"><img alt=\"\" height=\"145\" src=\"/media/article_images/2023/58/26/178-1651419347-Figure7.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 7. </strong>Gel electrophoresis of rs<strong><em>2236338 </em></strong>gene polymorphism amplified with specific pair of primers using conventional PCR. The PCR product was stained with ethidium bromide. The fragment length was 400 bp.</figcaption>\r\n</figure>\r\n</div>\r\n\r\n<div id=\"figure8\">\r\n<figure class=\"image\"><img alt=\"\" height=\"138\" src=\"/media/article_images/2023/58/26/178-1651419347-Figure8.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 8.</strong> Genotyping of <em>rs2236338</em> gene polymorphism after digestion with BsmF I and stained with ethidium bromide. Lanes 1 and 4: GG genotype lanes 2,5,6,8,10, and 11: AA, lanes 3,7,9, and 12: AG genotype; M: 100 bp DNA marker.</figcaption>\r\n</figure>\r\n</div>\r\n\r\n<div id=\"figure9\">\r\n<figure class=\"image\"><img alt=\"\" height=\"144\" src=\"/media/article_images/2023/58/26/178-1651419347-Figure9.jpg\" width=\"500\" />\r\n<figcaption><strong>Figure 9. </strong>Sequence analysis of the <em>rs2236338</em>, the forward strand. The A in the upper frame represents homozygous wild type (AA), the G in the second line of the frame represent the mutant genotype (GG), while the R in lower line represents the heterozygous genotype (AG).</figcaption>\r\n</figure>\r\n</div>\r\n\r\n<div id=\"Table-6\">\r\n<p><a href=\"https://jabet.bsmiab.org/table/178-1651419347-table6/\">Table-6</a><strong>Table 6.</strong> The occurrence of various genotypes and alleles of rs2236338 in hepatitis patients with and without HCC.</p>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Haplotype analysis</strong><br />\r\nThe SHEsis program was used to build haplotype blocks since the three polymorphisms are situated on the same gene.<a href=\"#Table-7\"> Table 7</a> shows the most frequent haplotypes in patients with and without HCC. The haplotype block TTA (T allele of rs7144366, T allele of rs8192917, and A allele of rs2236338) among patients without HCC had a higher prevalence than in HCC patients. (43.33% vs. 22.5%) with highly significant difference (OR= 0.38, 95%CI= 0.19-0.74, p= 0.005). in contrast, the haplotype block CCG was more common among patients with HCC (26.25%) than those without HCC (12.22%) with a significant difference (OR= 2.56, 95%= 1.14-5.71, p= 0.022). Although the haplotype block TTG was more common in patients with HCC (22.5%) than those without HCC (12.22%), the difference was not significant.</p>\r\n\r\n<div id=\"Table-7\">\r\n<p><a href=\"https://jabet.bsmiab.org/table/178-1651419347-table7/\">Table-7</a><strong>Table 7. </strong>The most frequent haplotype blocks.</p>\r\n\r\n<p> </p>\r\n</div>\r\n\r\n<p><strong>Linkage disequilibrium</strong><br />\r\nThe results of LD analysis are presented in <a href=\"#figure10\">Figure 10</a>. LD plot was constructed using combined genotype data from all patients. The SNP rs8192917 is in tight link with rs2236338 and rs8192917 (the measure D’ was 0.86). Furthermore, there was weak LD between rs8192917 and rs7144366 (the measure D’ was 0.57).</p>\r\n\r\n<div id=\"figure10\">\r\n<figure class=\"image\"><img alt=\"\" height=\"349\" src=\"/media/article_images/2023/58/26/178-1651419347-Figure10.jpg\" width=\"411\" />\r\n<figcaption><strong>Figure 10.</strong> Linkage disequilibrium pattern between different SNPs in GzmB gene. The red region shows strong linkage disequilibrium. D’ has a maximum value of 1.</figcaption>\r\n</figure>\r\n</div>"
},
{
"section_number": 4,
"section_title": "DISCUSSION",
"body": "<p>According to the result of the present study, the CC genotype of the SNP rs8192917 was significantly correlated with the development of HCC in patients with hepatitis B viral infection (OR= 3.93, 95%CI=1.13-13.62, p=0.031). This indicates that HBV patients with the CC genotype of this polymorphism are approximately four times more likely to develop HCC than individuals with the TT genotype. Furthermore, patients carrying the C allele of this polymorphism are at a 2.36-time higher risk than those with the T allele. On the other hand, the other two SNPs demonstrated no significant connection with the development of HCC in patients with HBV infection. Among available literature, there was no previous study that evaluated GzmB gene polymorphisms with the development of HCC. Alternatively, several studies have explored this association of these polymorphisms with other disorders and malignancies. In a recent Chinese study, Xu <em>et al</em>. [<a href=\"#r-8\">8</a>] recruited 973 patients with vitiligo and 2,147 age- and gender-matched family-unrelated controls. It was discovered that having the rs8192917 C allele increased the likelihood of having vitiligo by around 40 %. The significant relationship of this SNP was identified in all three genetic models (co-dominance, dominance, and recessive).<br />\r\nIn another study, Gaafar <em>et al.</em> investigated the role of GzmB gene polymorphism among 30 women with breast cancer and 38 other healthy Saudi Arabian women [<a href=\"#r-9\">9</a>]. The study indicated that women carrying the C allele of rs8192917 polymorphism are more susceptible to the disease compared with those carrying the T allele. In a Dutch study, Hurkmans <em>et al</em>. investigated the role of rs8192917 in 347 patients with stage IV lung cancer under anti-programmed cell death (PD-1) therapy [<a href=\"#r-10\">10</a>]. Patients with homozygous and heterozygous genotypes (TC and CC) had worse overall survival (OR: 1.60; 95%CI: 1.01 -2.52; p=0.044) and worse progression free survival (hazard ratio: 1.38; 95%CI:1.02 to 1.87; p=0.036) than those with wild types. In individuals infected with measles, rs8192917 was linked to subacute sclerosing panencephalitis, according to Yentur and his colleagues [<a href=\"#r-11\">11</a>]. Corrales’s working team suggested that rs8192917 homozygous TT has linkage with improved kidney allograft outcomes [<a href=\"#r-12\">12</a>]. In contrast, Espinoza <em>et al</em>. found that the CC genotype of rs8192917 in the patients who underwent bone marrow transplant was significantly associated with increased overall survival and reduced transplant-related mortality [<a href=\"#r-13\">13</a>]. Several mechanisms have been postulated to explain this effect of rs8192917 as a risk for many diseases. This polymorphism is located in exon 2 of the gene. It involves a substitution of thymine with cytosine with a subsequent substitution of glutamine with arginine at amino acid number 55 of the polypeptide chain.<br />\r\nThe most plausible explanation was proposed by McIlory <em>et al.</em> [<a href=\"#r-14\">14</a>], who transfected primary glioblastoma cells with a mutant allele of this polymorphism and used x-ray crystallography to study the interaction between different amino acids in the GzmB structure. The study revealed that the presence of glutamine 55 in this peptide forms a hydrogen bond with the amino acid methionine 242. This interaction anchors the C-terminal α-helix to the surface of the protein, which seems to have a functional role in the catalytic properties of the enzyme. However, this interaction is disrupted when glutamine is substituted by arginine, and the protein loses some of its activity. In the same context, Oboshi <em>et al. </em>[<a href=\"#r-15\">15</a>] have shown that rs8192917 can affect NK cell cytotoxicity. However, the authors did not explain the mechanism by which this polymorphism affects NK cells. These findings suggest that this polymorphism influences cytotoxic T cell function by altering the protein binding capacity with its substrate. In the present study, the haplotype CCG (C allele of rs7144366, C allele of rs8192917, and G allele of rs2236338) was significantly associated with an increased risk of HCC in patients with CHB. These alleles correspond to alanine, arginine, and histidine amino acid, respectively. This is in accordance with several previous reports that have investigated the three SNPs in different diseases. In a Korean study, Jeong <em>et al.</em> [<a href=\"#r-16\">16</a>] investigated a total of 249 patients with non-segmented vitiligo (NSV) and 455 healthy controls to determine the impact of 5 SNPs (rs2236337, rs2236338, rs11539752, rs10909625, and rs8192917) in GzmB gene on patients’ susceptibility to this autoimmune disease. The haplotypes CGCCC consisting of rs2236337, rs2236338, rs11539752, rs10909625, and rs8192917 demonstrated a significant association with the disease. In another study, individuals presenting mutant haplotype of the three SNPs (rs7144366, rs8192917, and rs2236338) were 16 times more likely to have breast cancer as compared with those with wild type haplotype [<a href=\"#r-9\">9</a>].<br />\r\nThe mechanism by which the CCG haplotype increases the risk of HCC is not fully understood. Both mutant and wild type haplotypes of the alternative GzmB polypeptide isoforms are thought to have equal expression, stability, and proteolytic activity [<a href=\"#r-14\">14</a>]. However, Gaafar et al. [<a href=\"#r-9\">9</a>] found that CCG induced less apoptosis than TAA GzmB, while Sun <em>et al. </em>[<a href=\"#r-17\">17</a>] found no difference. Other studies reported that the triple-mutated granzyme B variant encodes three amino acid changes with altered biological functions [<a href=\"#r-18\">18</a>]. Alternatively, McIlory et al. [<a href=\"#r-14\">14</a>] claimed that the three altered amino acids did not affect the active site or the substrate-binding cleft configuration. Furthermore, none of the three locations are involved in the active site or substrate-binding pocket. Additionally, the location of the triple mutant’s side chains revealed no steric incompatibility between the mutant side chains and the wild-type conformation. Apoptosis induction, in contrast, differed significantly between RAH and QPY GzmB. GzmB QPY (for glutamine, proline, and tyrosine, respectively) triggered apoptosis, as shown by the fact that caspase-3 is active in cell lysates and that dead cells let out LDH. But RAH GzmB (RAH stands for arginine, alanine, and histidine) did not cause caspase activity or cell death more than what was seen in control transfected cells. Three separate cell lines from different patients all showed the same results [<a href=\"#r-14\">14</a>].<br />\r\nIn the present study, The SNP rs8192917 is in tight link with rs2236338 (the measure D’ was 0.86). Furthermore, there was weak LD between rs2236338 and rs7144366 (the measure D’ was 0.57). Most previous studies indicated the presence of such a link, although with different degrees.<br />\r\nThere is a significant degree of linkage disequilibrium between these mutations, as shown by many lines of evidence. Patients who are heterozygous for the Y245H variation are invariably heterozygous for the Q48R and P88A variants, while patients who were homozygous for the H245 variant were also homozygous for the R48 and A88 variants. Furthermore, cloning and sequencing of PCR products spanning exons 1–5 of the GzmB gene from a single heterozygous at all three sites confirmed that one chromosome coded for Q48 P88 Y245, whereas the other coded for R48 A88 H245 [<a href=\"#r-14\">14</a>]. A study in Europe discovered a high level of LD among the three non-synonymous SNPs (rs8192917, rs11539752, and rs2236338) among European populations, resulting in different proteins [<a href=\"#r-19\">19,20</a>]. Jin <em>et al.</em> [<a href=\"#r-21\">21</a>] showed that rs8192917C was in a very strong LD with two other common non-synonymous SNPs, rs11539752 (r<sup>2</sup> = 0.99) and rs2236338 (r<sup>2</sup> = 0.93). In another study, the three SNPs had strong LD. In particular, rs8192917 had a stronger LD with rs11539752 (D′ = 0.951, r<sup>2</sup> = 0.861) and rs2236338 (D′ = 0.942, r<sup>2</sup> = 0.464,). A strong LD was also observed between rs8192917 and rs11539752 (D′ = 1.000, r<sup>2</sup> = 0.549) [<a href=\"#r-15\">15</a>].</p>"
},
{
"section_number": 5,
"section_title": "CONCLUSIONS",
"body": "<p>Collectively, these data indicate that allele C of this SNP could be considered a risk factor for developing HCC. Furthermore, the haplotype block CCG (corresponding to the TC allele of rs7144366, the C allele of rs8192917, and the G allele of rs2236338) could also be considered a risk factor for the development of HCC in patients with CHB. In contrast, the haplotype block TAA of these SNPs has a protective role. There is a linkage disequilibrium between the three SNPs, especially between rs2236338 and rs8192917.</p>"
},
{
"section_number": 6,
"section_title": "ACKNOWLEDGEMENT",
"body": "<p>The authors highly appreciate the great cooperation of included patients to complete the study. Also, we acknowledge the great help from all staff at Medical Research Unit/ College of Medicine/ Al-Nahrain University for their effort in molecular assays.</p>"
},
{
"section_number": 7,
"section_title": "AUTHOR CONTRIBUTIONS",
"body": "<p>QSA conceptualized and supervised the experiment. QSA and HSA performed the experiment. HAS and FAA assisted in the sample collection and gross data recording and provided research assistance for data collection. QSA and HSA analyzed the data and interpreted the results. QSA, FAA and HAS drafted the manuscript and edited the manuscript. All authors agreed to be responsible for all elements of the work, including its accuracy and integrity.</p>"
},
{
"section_number": 8,
"section_title": "CONFLICTS OF INTEREST",
"body": "<p>There is no conflict of interest among the authors.</p>"
}
],
"figures": [
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/58/26/178-1651419347-Figure1.jpg",
"caption": "Figure 1. Gel electrophoresis (1% agarose) of rs7144366 gene polymorphism amplified with specific pair of primers using conventional PCR run for 45 min at 129 V. The PCR product was stained with ethidium bromide. The fragment length was 97bp.",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/58/26/178-1651419347-Figure2.jpg",
"caption": "Figure 2. Genotyping of rs7144366 gene polymorphism after digestion with SfcI and stained with ethidium bromide. Lanes 1,3,6,7,8, and 11: TT; lanes 2,5, and 10: CT, lanes 4 and 9: CC genotype; M: DNA 50 bp DNA marker.",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/58/26/178-1651419347-Figure3.jpg",
"caption": "Figure 3. Sequence analysis of the rs7144366, the reverse strand. The R in the upper frame represents the heterozygous genotype (TC), the G in the middle frame represents the mutant genotype (CC), and the A in the bottom frame represents the homozygous wild type of genotype (TT).",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/58/26/178-1651419347-Figure4.jpg",
"caption": "Figure 4. Gel electrophoresis (1% agarose) of rs 8192917 gene polymorphism amplified with specific pair of primers using conventional PCR run for 45 min at 129 V. The PCR product was stained with ethidium bromide. The fragment length was 420 bp.",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/58/26/178-1651419347-Figure5.jpg",
"caption": "Figure 5. Genotyping of rs7144366 gene polymorphism after digestion with BsmA I and stained with ethidium bromide. Lanes 1,2,4,7,8,10,11, 13 and 14: TT; lanes 5,6 and 12: CT, lanes 4 and 9: CC genotype; M: 100 bp DNA maker.",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/58/26/178-1651419347-Figure6.jpg",
"caption": "Figure 6. Sequence analysis of the rs8192917, the reverse strand. The homozygous wild type is represented by the letter A in the top frame (TT), the G in the second line of the frame represents the mutant genotype (CC), while the R in the lower line represents the heterozygous genotype (TC).",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/58/26/178-1651419347-Figure7.jpg",
"caption": "Figure 7. Gel electrophoresis of rs2236338 gene polymorphism amplified with specific pair of primers using conventional PCR. The PCR product was stained with ethidium bromide. The fragment length was 400 bp.",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/58/26/178-1651419347-Figure8.jpg",
"caption": "Figure 8. Genotyping of rs2236338 gene polymorphism after digestion with BsmF I and stained with ethidium bromide. Lanes 1 and 4: GG genotype lanes 2,5,6,8,10, and 11: AA, lanes 3,7,9, and 12: AG genotype; M: 100 bp DNA marker.",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/58/26/178-1651419347-Figure9.jpg",
"caption": "Figure 9. Sequence analysis of the rs2236338, the forward strand. The A in the upper frame represents homozygous wild type (AA), the G in the second line of the frame represent the mutant genotype (GG), while the R in lower line represents the heterozygous genotype (AG).",
"featured": false
},
{
"figure": "https://jabet.bsmiab.org/media/article_images/2023/58/26/178-1651419347-Figure10.jpg",
"caption": "Figure 10. Linkage disequilibrium pattern between different SNPs in GzmB gene. The red region shows strong linkage disequilibrium. D’ has a maximum value of 1.",
"featured": false
}
],
"authors": [
{
"id": 469,
"affiliation": [
{
"affiliation": "Department of Chemistry and Biochemistry, College of Medicine, Al-Nahrain University, Iraq"
}
],
"first_name": "Hussein S.",
"family_name": "Alshamary",
"email": null,
"author_order": 1,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 116
},
{
"id": 470,
"affiliation": [
{
"affiliation": "Medical Research Unit, College of Medicine, Al-Nahrain University, Iraq"
}
],
"first_name": "Qasim S. Al",
"family_name": "Mayah",
"email": "qasimsharhan@gmail.com",
"author_order": 2,
"ORCID": null,
"corresponding": true,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "Qasim S. Al-Mayah, PhD; Medical Research Unit, College of Medicine, Al-Nahrain University, Iraq, e-mail: qasimsharhan@gmail.com",
"article": 116
},
{
"id": 471,
"affiliation": [
{
"affiliation": "Department of Internal Medicine, College of Medicine, Al-Nahrain University, Iraq"
}
],
"first_name": "Fadhil Abdulla Abdul",
"family_name": "Ridha",
"email": null,
"author_order": 3,
"ORCID": null,
"corresponding": false,
"co_first_author": false,
"co_author": false,
"corresponding_author_info": "",
"article": 116
}
],
"views": 744,
"downloads": 134,
"references": [
{
"id": 3671,
"serial_number": 1,
"pmc": null,
"reference": "Ferlay J, Shin HR, Bray F, Forman D, Mathers C, Parkin DM. Estimates of worldwide burden of cancer in 2008: GLOBOCAN 2008. Int J Cancer. 2010 Dec 15;127(12):2893-917.",
"DOI": null,
"article": 116
},
{
"id": 3672,
"serial_number": 2,
"pmc": null,
"reference": "Bertoletti A, Tan AT. HBV as a target for CAR or TCR-T cell therapy. Curr Opin Immunol. 2020 Oct; 66:35-41.",
"DOI": null,
"article": 116
},
{
"id": 3673,
"serial_number": 3,
"pmc": null,
"reference": "Tan A, Koh S, Bertoletti A. Immune response in hepatitis B virus infection. Cold Spring Harb Perspect Med. 2015;5(8): a021428.",
"DOI": null,
"article": 116
},
{
"id": 3674,
"serial_number": 4,
"pmc": null,
"reference": "Lieberman J. Granzyme A activates another way to die. Immunol Rev. 2010;235(1):93-104.",
"DOI": null,
"article": 116
},
{
"id": 3675,
"serial_number": 5,
"pmc": null,
"reference": "Ben Safta T, Ziani L, Favre L, Lamendour L, Gros G, Mami-Chouaib F, et al. Granzyme B-activated p53 interacts with Bcl-2 to promote cytotoxic lymphocyte-mediated apoptosis. J Immunol. 2015 Jan 1;194(1):418-28.",
"DOI": null,
"article": 116
},
{
"id": 3676,
"serial_number": 6,
"pmc": null,
"reference": "Girnita DM, Webber SA, Brooks MM, Ferrell R, Girnita AL, Burckart GJ, et al. Genotypic variation and phenotypic characterization of granzyme B gene polymorphisms. Transplantation. 2009 Jun;87(12):1801-1806.",
"DOI": null,
"article": 116
},
{
"id": 3677,
"serial_number": 7,
"pmc": null,
"reference": "Mhaidat NM, Al-azzam SI, Alzoubi KH, Khabour OF, Gharaibeh BF. Granzyme B gene polymorphisms, colorectal cancer risk, and metastasis. J Cancer Res Ther. 2014 Jul-Sep;10(3):587-90.",
"DOI": null,
"article": 116
},
{
"id": 3678,
"serial_number": 8,
"pmc": null,
"reference": "Xu M, Liu Y, Liu Y. Li X, Chen G, Dong W, et al. Genetic polymorphisms of GZMB and vitiligo: A genetic association study based on Chinese Han population. Sci Rep. 2018;8, 13001",
"DOI": null,
"article": 116
},
{
"id": 3679,
"serial_number": 9,
"pmc": null,
"reference": "Gaafar A, Aljurf MD, Al Sulaiman A, Iqniebi A, Manogaran PS, Mohamed GE, et al. Defective gammadelta T cell function and granzyme B gene polymorphism in a cohort of newly diagnosed breast cancer patients. Exp Hematol. 2009; 37:838 48.",
"DOI": null,
"article": 116
},
{
"id": 3680,
"serial_number": 10,
"pmc": null,
"reference": "Hurkmans DP, Basak EA, Schepers N, Oomen-De Hoop E, Van der Leest CH, El Bouazzaoui S, et al. Granzyme B is correlated with clinical outcome after PD-1 blockade in patients with stage IV non-small-cell lung cancer. J Immunother Cancer. 2020 May;8(1): e000586.",
"DOI": null,
"article": 116
},
{
"id": 3681,
"serial_number": 11,
"pmc": null,
"reference": "Yentur SP, Aydin HN, Gurses C, Demirbilek V, Kuru U, Uysal S, et al. Granzyme B gene polymorphism associated with subacute sclerosing panencephalitis. Neuropediatrics. 2014;45, 309–313.",
"DOI": null,
"article": 116
},
{
"id": 3682,
"serial_number": 12,
"pmc": null,
"reference": "Corrales-Tellez E, Vu D, Shah T, Hutchinson I, Min DI. Association between granzyme B and perforin I polymorphisms and allograft outcomes in Hispanic kidney transplant recipients. Clin. Transpl. 2013;27: E308–E315.",
"DOI": null,
"article": 116
},
{
"id": 3683,
"serial_number": 13,
"pmc": null,
"reference": "Espinoza LJ, Takami A, Nakata K, Yamada K, Onizuka M, Kawase T, et al. Genetic variants of human granzyme B predict transplant outcomes after HLA matched unrelated bone marrow transplantation for myeloid malignancies. PLoS ONE. 2011:6: e23827",
"DOI": null,
"article": 116
},
{
"id": 3684,
"serial_number": 14,
"pmc": null,
"reference": "McIlroy D, Cartron PF, Tuffery P, Dudoit Y, Samri A, Autran B, et al. A triple mutated allele of granzyme B incapable of inducing apoptosis. Proc Natl Acad Sci USA. 2003; 100:2562 7.",
"DOI": null,
"article": 116
},
{
"id": 3685,
"serial_number": 15,
"pmc": null,
"reference": "Oboshi W, Watanabe T, Hayashi K, Nakamura T, Yukimasa N. QPY/RAH haplotypes of the GZMB gene are associated with natural killer cell cytotoxicity. Immunogenetics. 2018;70, 29–36.",
"DOI": null,
"article": 116
},
{
"id": 3686,
"serial_number": 16,
"pmc": null,
"reference": "Jeong KH, Kim SK, Seo JK, Shin MK, Lee MH. Association of GZMB polymorphisms and susceptibility to non-segmental vitiligo in a Korean population. Sci Rep. 2021;11(1):397.",
"DOI": null,
"article": 116
},
{
"id": 3687,
"serial_number": 17,
"pmc": null,
"reference": "Sun J, Bird CH, Thia KY, Matthews AY, Trapani JA, Bird PI. Granzyme B encoded by the commonly occurring human RAH allele retains pro-apoptotic activity. J Biol Chem. 2004; 279:16907-16911.",
"DOI": null,
"article": 116
},
{
"id": 3688,
"serial_number": 18,
"pmc": null,
"reference": "Zaitsu M, Yamamoto K, Ishii E, Teramura T, Nakadate N, Sako M, et al. High frequency of QPY allele and linkage disequilibrium of granzyme B in Epstein-Barr virus associated hemophagocytic lymphohistiocytosis. Tissue Antigens. 2004; 64: 611.",
"DOI": null,
"article": 116
},
{
"id": 3689,
"serial_number": 19,
"pmc": null,
"reference": "Guan, F, Niu Y, Zhang T, Liu S, Ma L, Qi T, et al. Two-stage association study to identify the genetic susceptibility of a novel common variant of rs2075290 in ZPR1 to type 2 diabetes. Sci Rep. 2016; 6,29586.",
"DOI": null,
"article": 116
},
{
"id": 3690,
"serial_number": 20,
"pmc": null,
"reference": "Guan F, Wei S, Zhang C, Zhang H, Zhang B, et al. A population-based association study of 2q32.3 and 8q21.3 loci with schizophrenia in Han Chinese. J Psychiatric Res. 2013; 47:712–717.",
"DOI": null,
"article": 116
},
{
"id": 3691,
"serial_number": 21,
"pmc": null,
"reference": "Jin Y, Birlea SA, Fain PR, Gowan K, Riccardi SL, Holland PJ, et al. Variant of TYR and autoimmunity susceptibility loci in generalized vitiligo. New Engl J Med. 2010; 362:1686–97.",
"DOI": null,
"article": 116
}
]
}
]
}
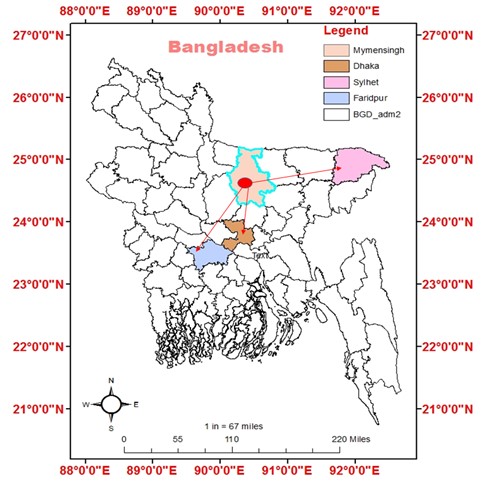{kind=link}
{kind=link}
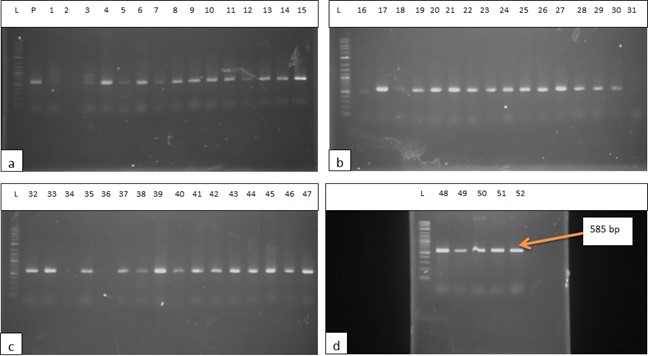{kind=link}
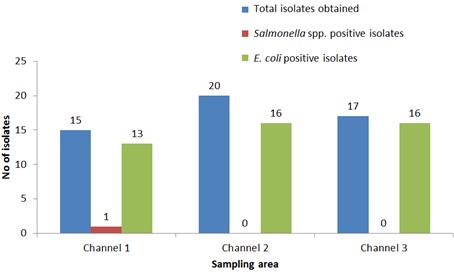{kind=link}
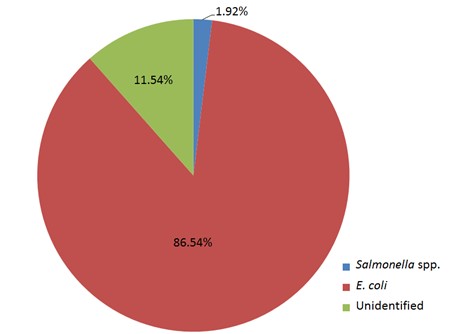{kind=link}
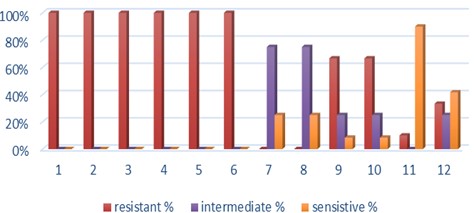{kind=link}
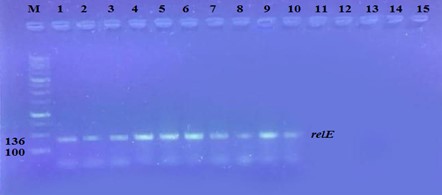{kind=link}
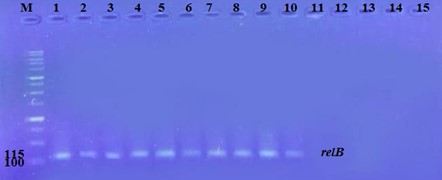{kind=link}
{kind=link}
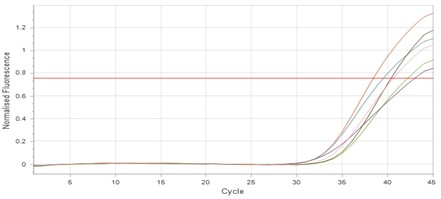{kind=link}
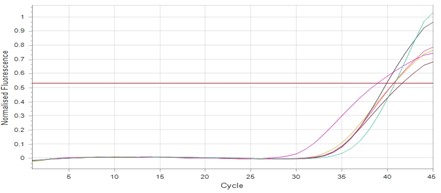{kind=link}
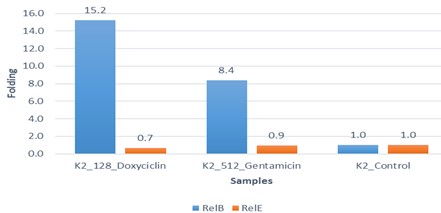{kind=link}
{kind=link}
{kind=link}
{kind=link}
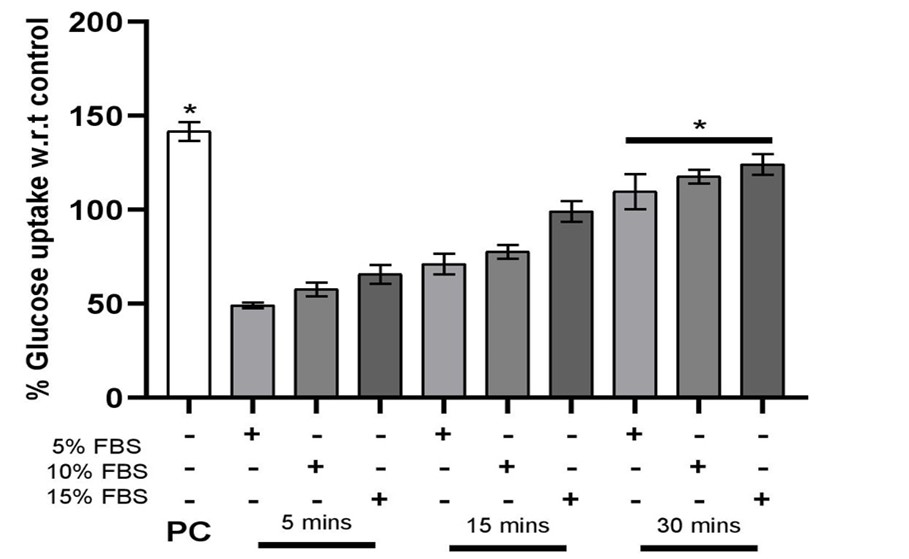{kind=link}
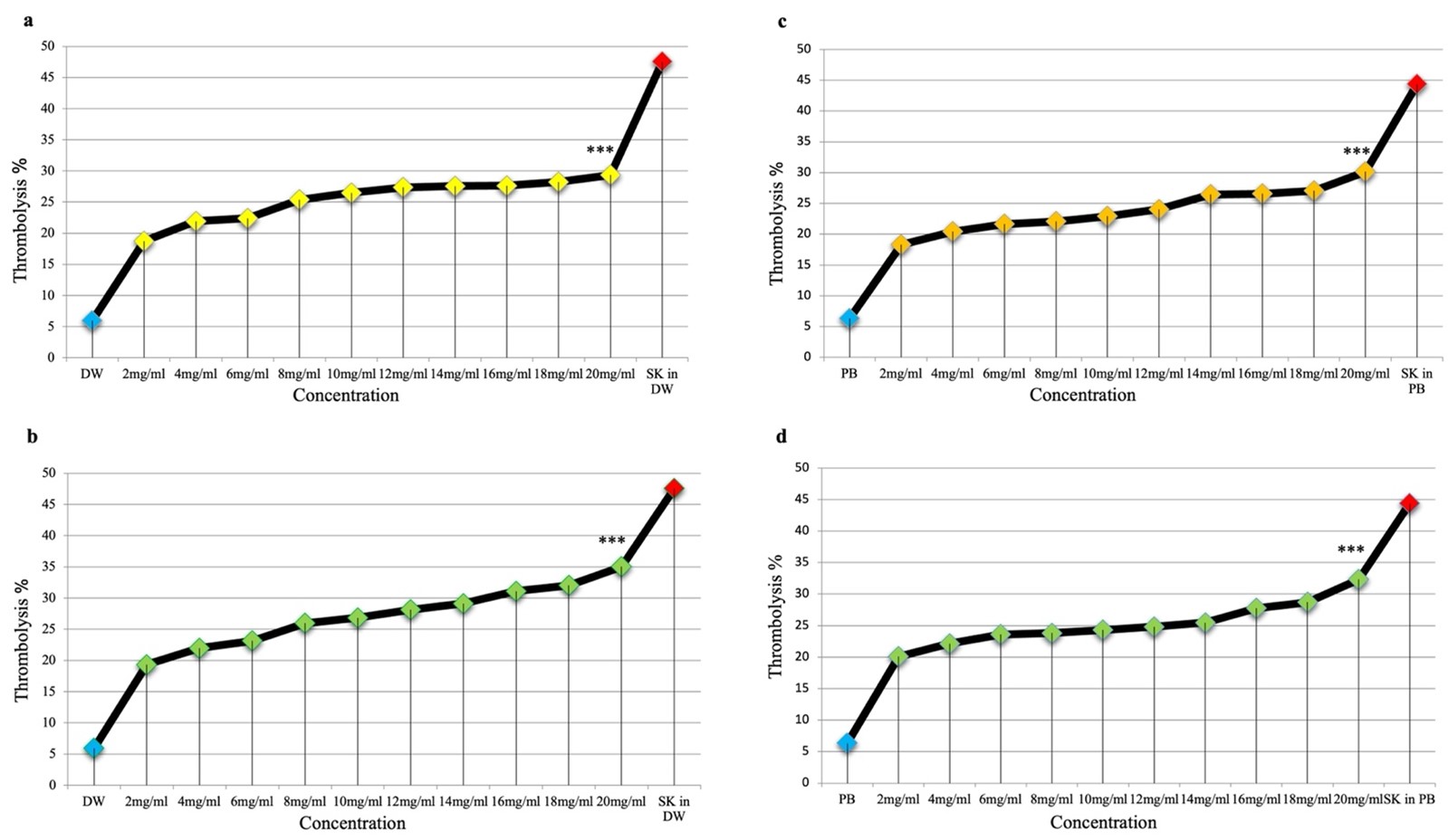{kind=link}
{kind=link}
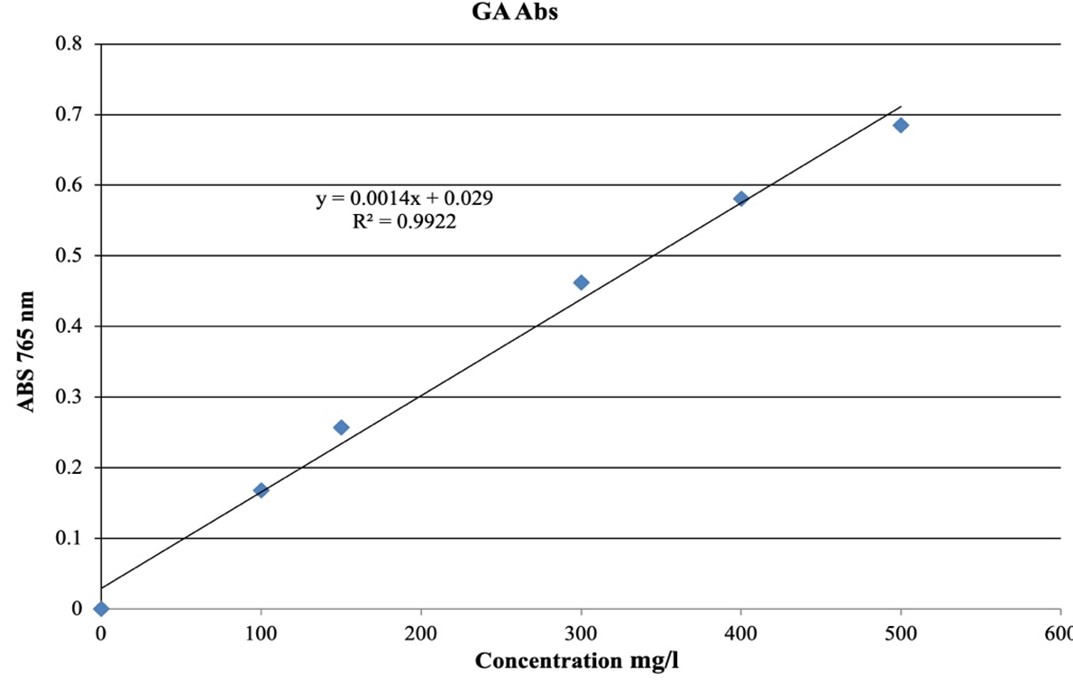{kind=link}
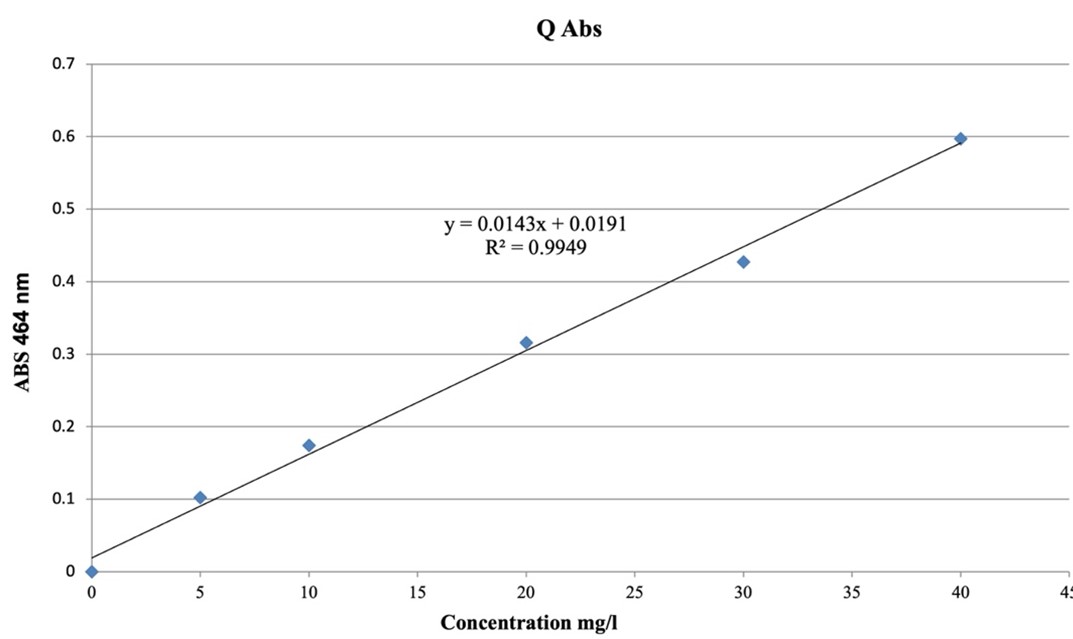{kind=link}
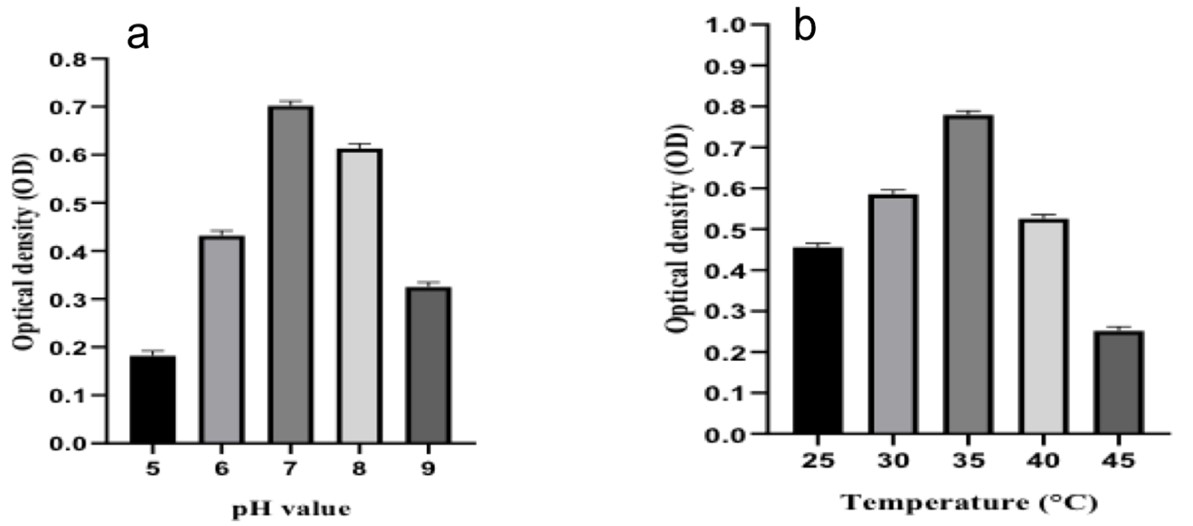{kind=link}
{kind=link}
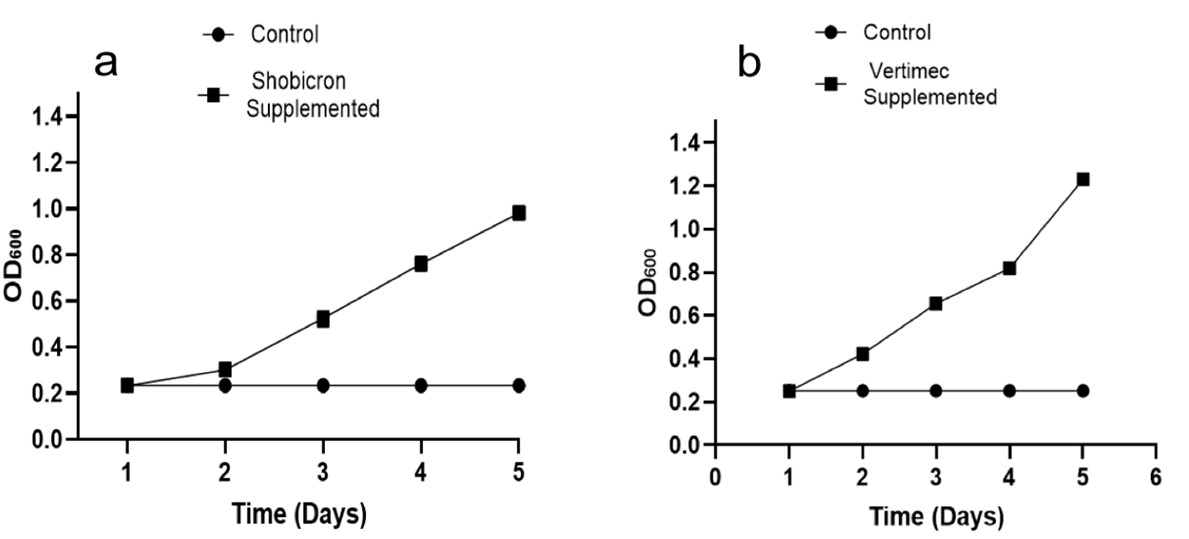{kind=link}
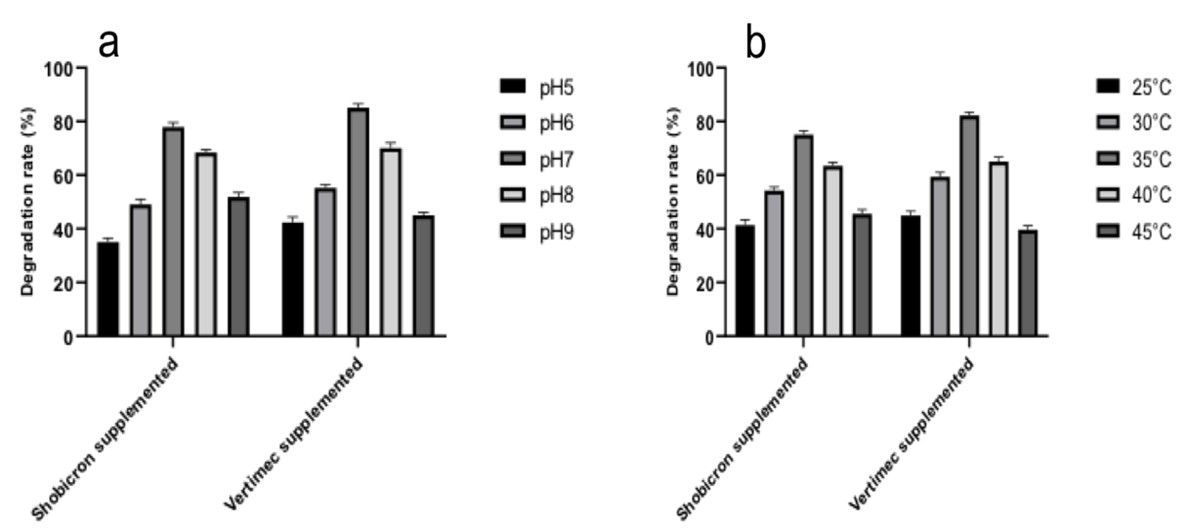{kind=link}
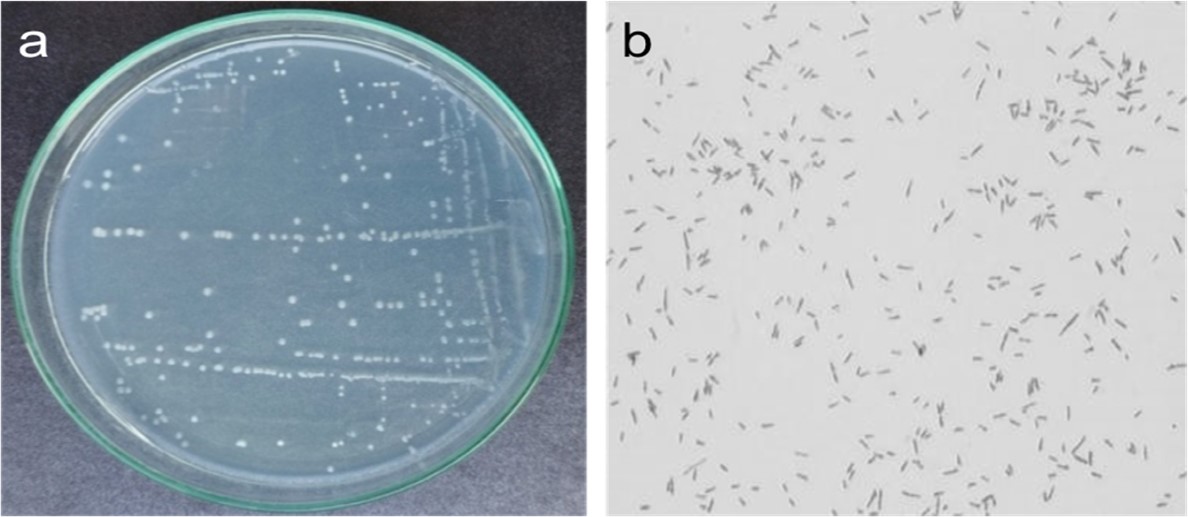{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
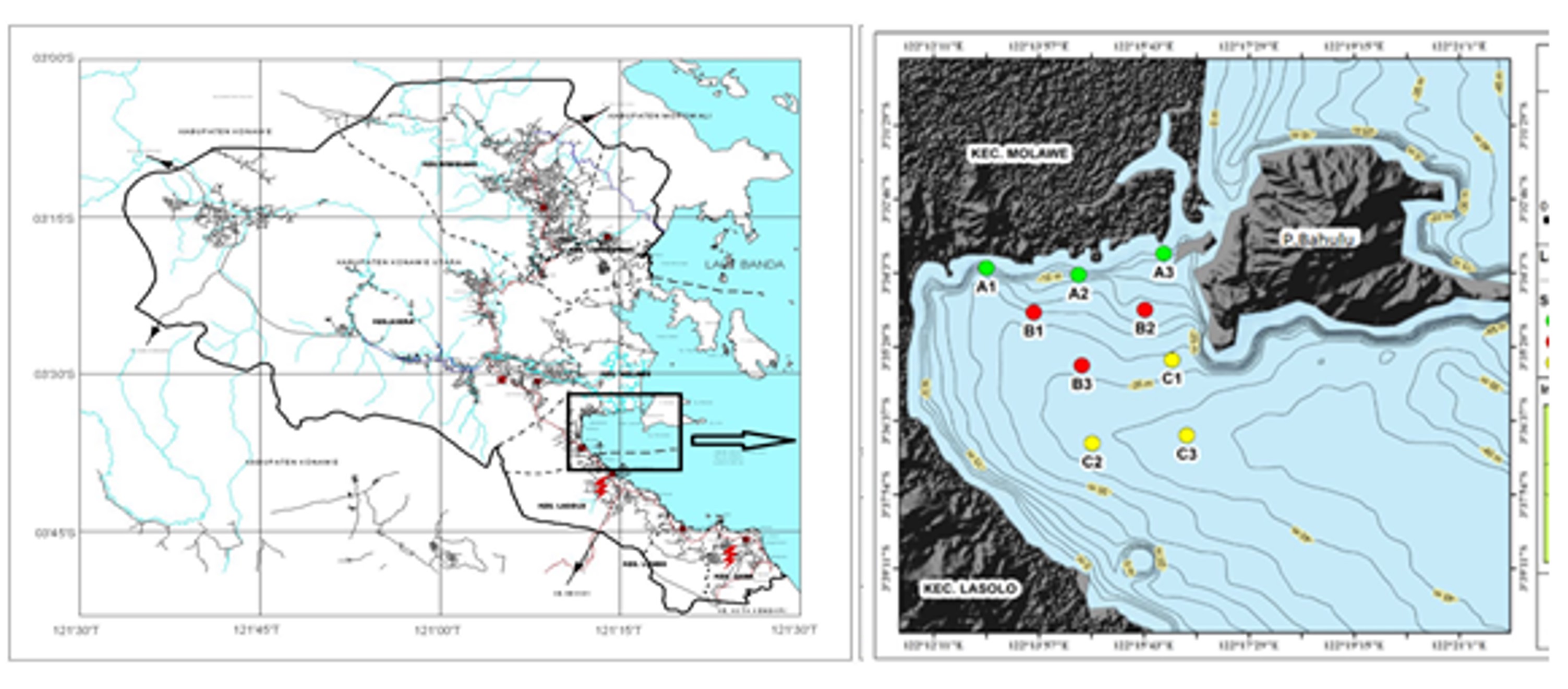{kind=link}
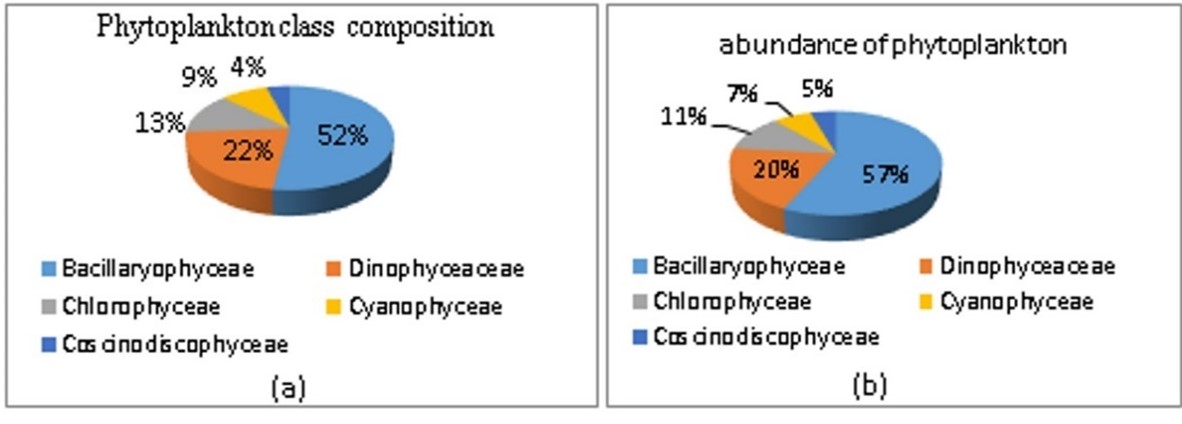{kind=link}
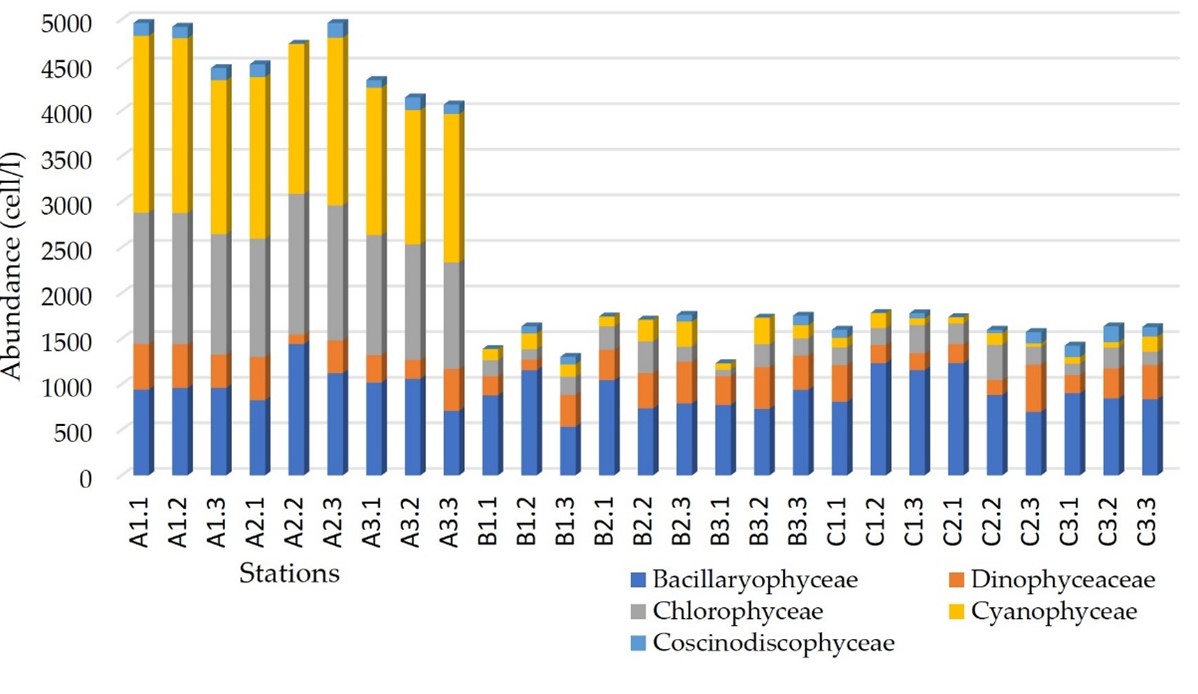{kind=link}
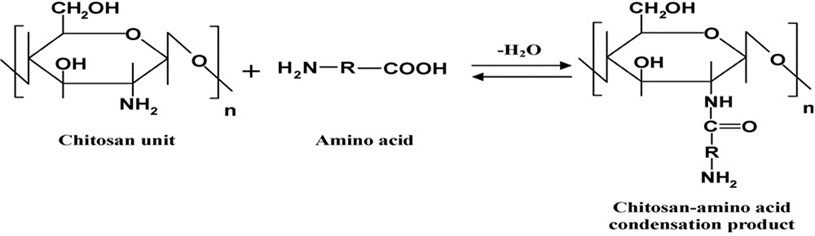{kind=link}
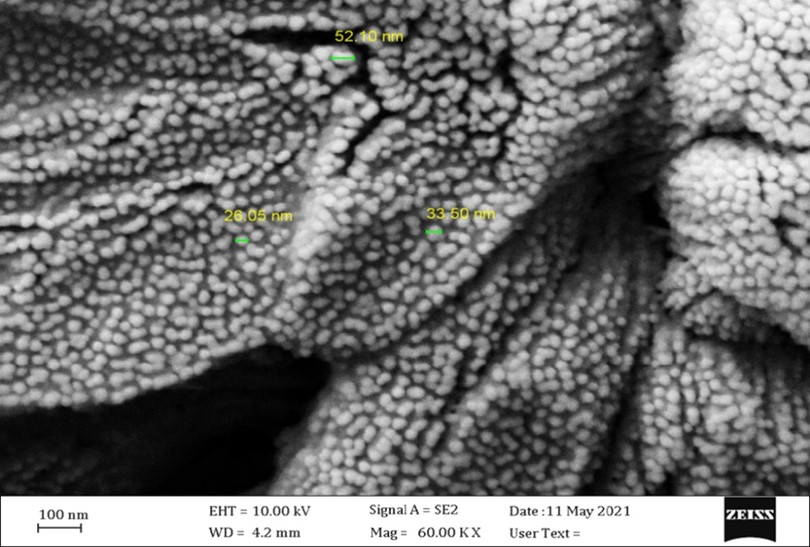{kind=link}
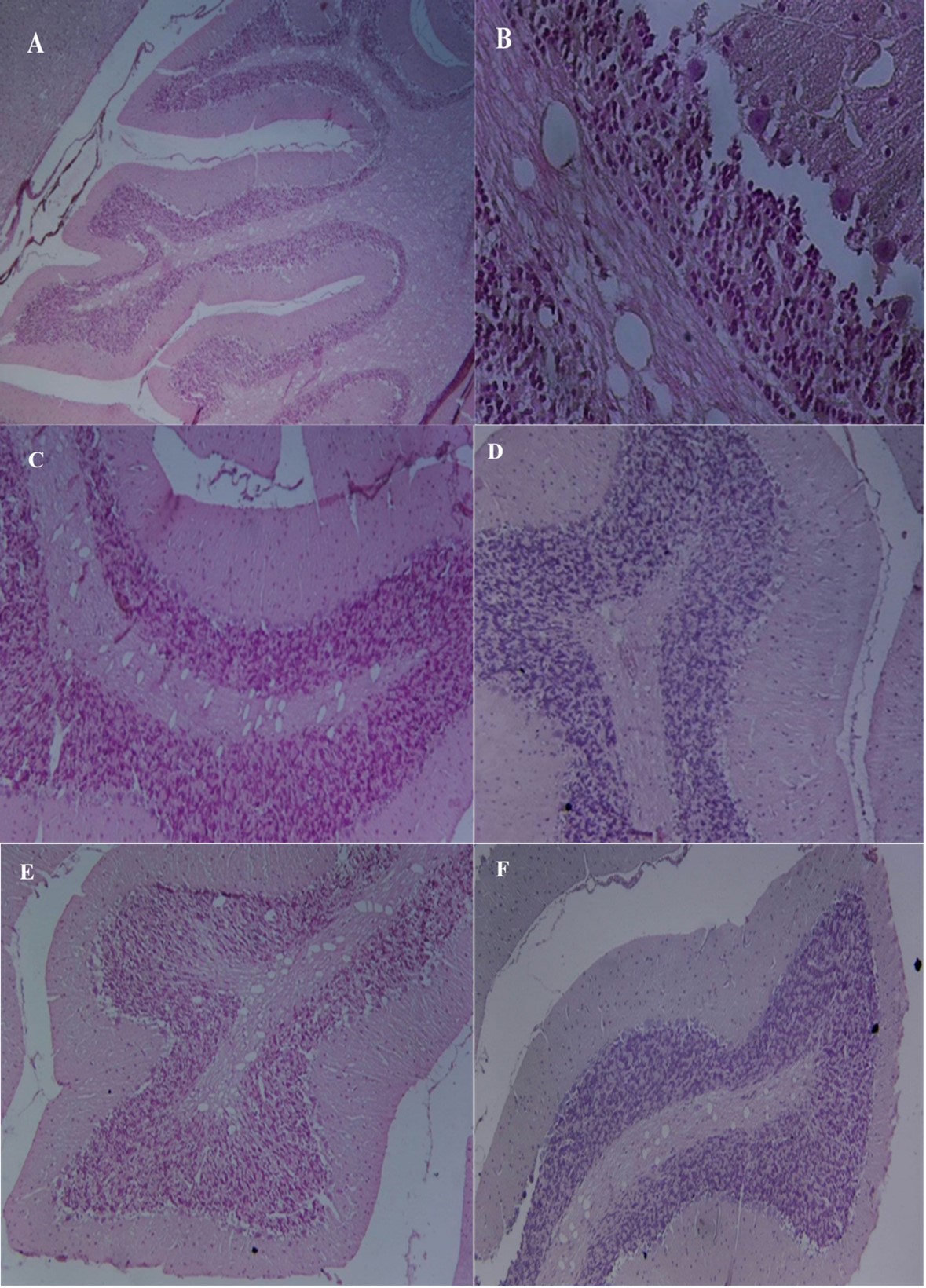{kind=link}
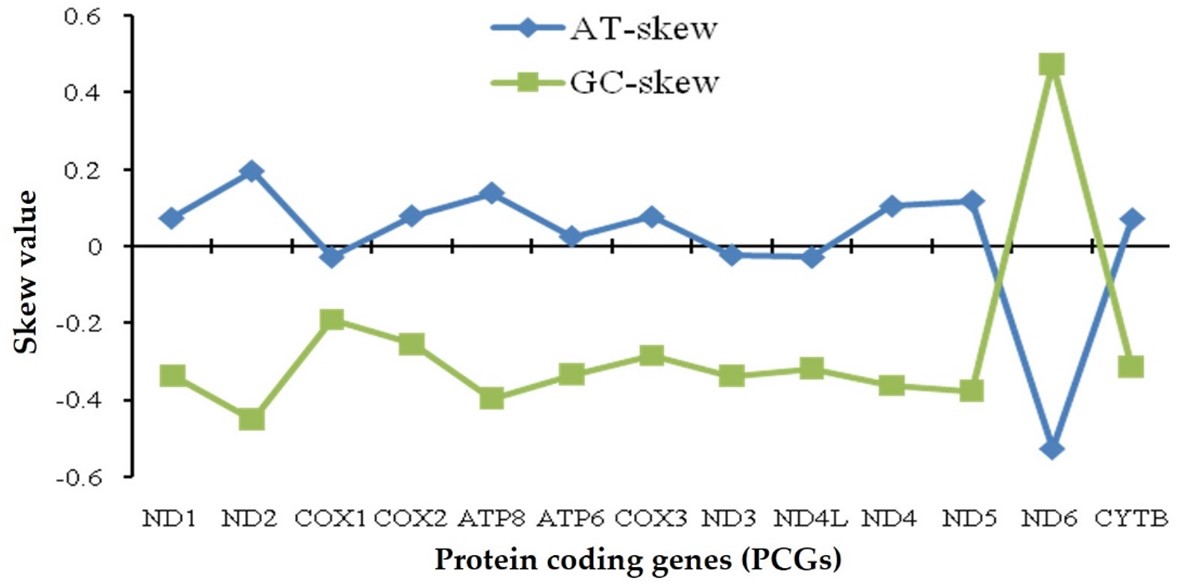{kind=link}
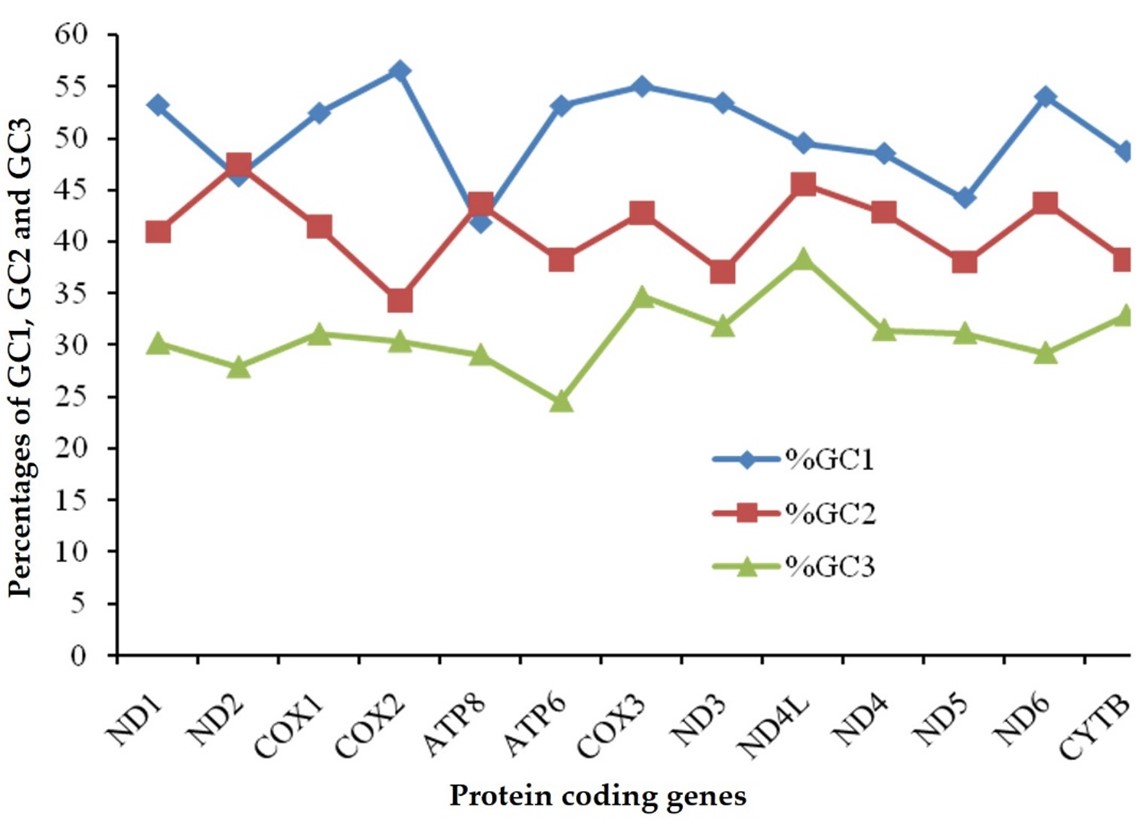{kind=link}
{kind=link}
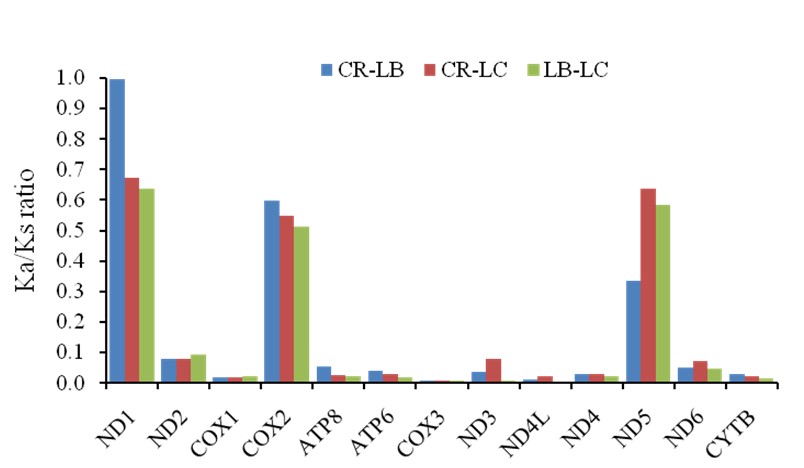{kind=link}
{kind=link}
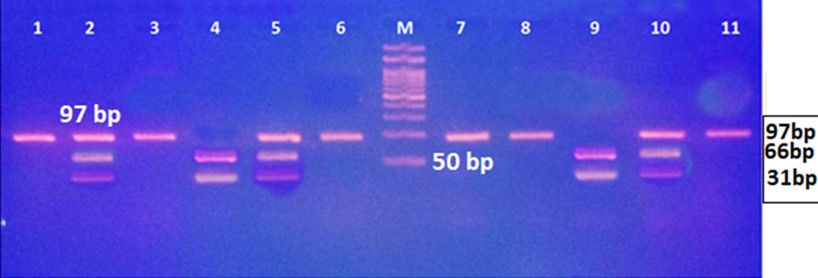{kind=link}
{kind=link}
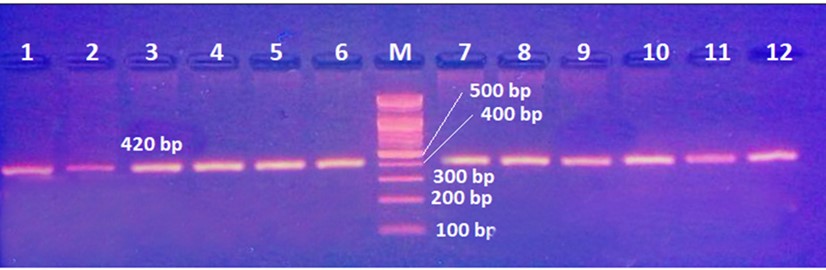{kind=link}
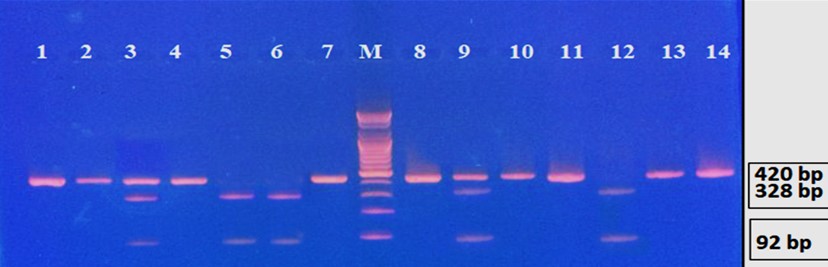{kind=link}
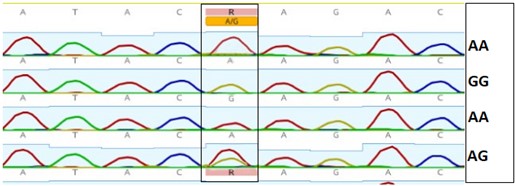{kind=link}
{kind=link}
{kind=link}
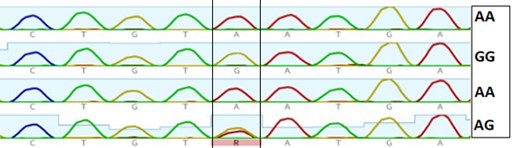{kind=link}
{kind=link}